
Complement System
补体是由朱尔斯·博尔代发现的,它是正常血浆中一种热不稳定的成分,能够调理并杀灭细菌 。补体系统指的是一系列在血液和组织液中循环的蛋白质。大多数蛋白质通常处于非活性状态,但当识别到微生物的分子成分时,它们会通过酶级联反应依次被激活——一种蛋白质的激活会酶促裂解并激活级联反应中的下一个蛋白质。补体可以通过三种不同的途径被激活( 图 1 ),每条途径都能激活 C3 ,将其裂解成一个大的片段 C3b (作为调理素 )和一个较小的片段 C3a (过敏毒素,促进炎症)。激活的 C3 可以触发溶菌途径 ,从而损伤细胞和某些细菌的质膜。该过程产生的 C5a 可吸引巨噬细胞和中性粒细胞,并激活肥大细胞 。
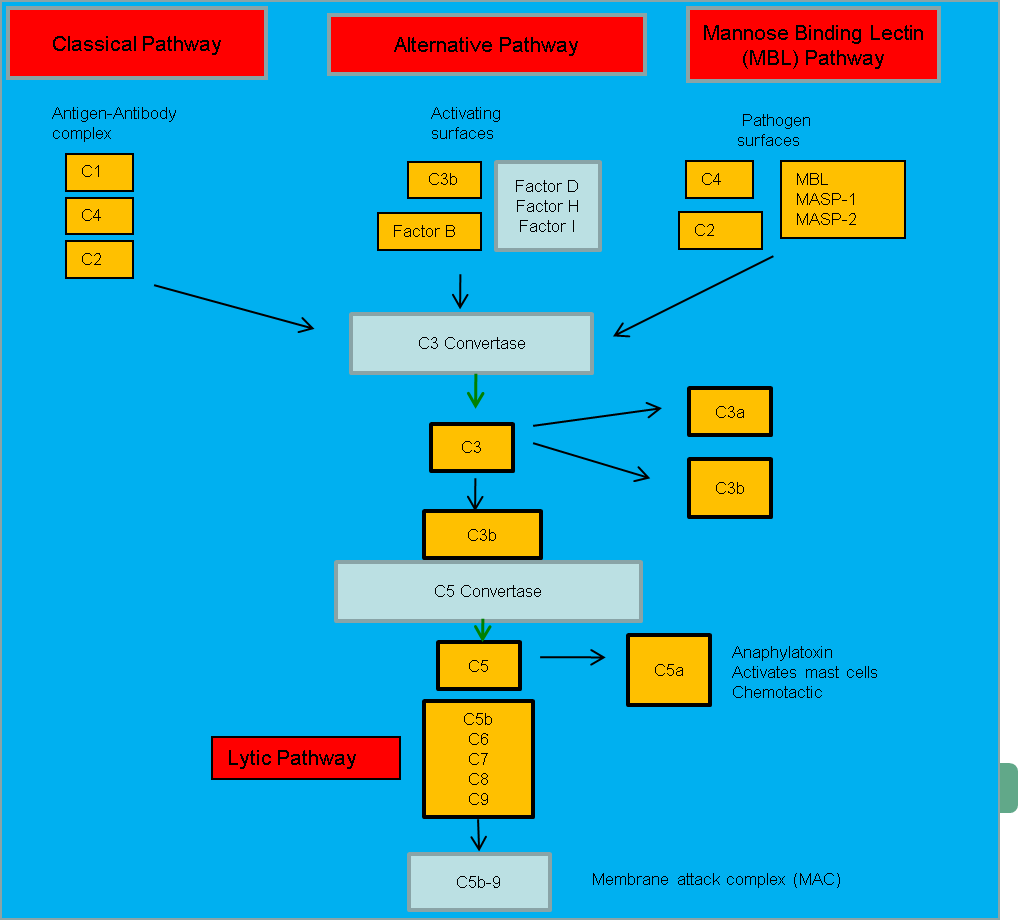
经典途径
该通路涉及补体成分 C1 、 C2 和 C4 。 抗体-抗原复合物与 C1 结合可触发该通路,C1 本身包含三个亚成分: C1q 、 C1r 和 C1s 。该通路生成 C3 转化酶 C4b2a ,后者可将 C3 裂解成两个片段;较大的片段 C3b 可共价结合到微生物病原体的表面并对其进行调理 ;较小的片段 C3a 可激活肥大细胞 ,导致组胺等血管活性介质的释放。
替代路径
该通路涉及多种因子 B、D、H 和 I ,这些因子彼此相互作用,并与 C3b 结合,形成 C3 转化酶 C3bBb ,后者能够激活更多的 C3,因此该通路有时被称为“放大环”。细菌和真菌细胞壁的存在会促进该环的激活,但正常哺乳动物细胞表面的分子会抑制该环的激活。
甘露糖结合凝集素途径
该通路通过甘露糖结合凝集素 ( MBL )与病原体表面甘露糖残基的结合而被激活。这反过来激活了 MBL 相关的丝氨酸蛋白酶 MASP-1 和 MASP-2 ,它们激活 C4 和 C2 ,形成 C3 转化酶 C4b2a 。
裂解途径
该途径由 C5 的裂解以及 C5b 与靶标的结合启动。C6 、C7、C8 和 C9 与 C5b 结合,形成膜攻击复合物 ( MAC )。当 MAC 插入某些细菌的外膜时,可导致细菌裂解死亡。表面结合抗体的红细胞也能激活经典途径和裂解途径,从而变得易受裂解。
补体在疾病中的作用
补体系统在炎症反应和抵御某些细菌感染中发挥着关键作用。在输血不相容反应以及伴随自身免疫性疾病的有害免疫反应中,补体也可能被激活。单个补体成分或系统抑制剂的缺乏可导致多种疾病( 表 1 ),这表明它们在抵御疾病方面发挥着一定作用。
表 1. 与补体缺陷相关的疾病
| 补体缺乏症 | 疾病 |
| C3 和因子 B | 严重细菌感染 |
| C3b-INA、C6 和 C8 | 严重的奈瑟菌感染 |
| 早期 C 组分 C1、C4、C2 的缺乏。 | 系统性红斑狼疮(SLE)、肾小球肾炎和多发性肌炎 |
| C1 抑制剂 | 遗传性血管性水肿 |
© 本作品的版权归作者所有。
Hits: 137
孟博士您好,我两周没回家,前两天回家发现地上有几颗粪便,分不清是老鼠的还是蝙蝠的,已经干了,很硬,没多想就直接用手拿了,我想问一下风干的老鼠或者蝙蝠的粪便会传播狂犬病吗?因为我打算把屋子里大扫除一下看看角落还没有粪便,我担心破皮的地方不小心碰到粪便。可以请孟博士拨冗解答一下我的疑惑吗?谢谢您!
想多了
孟哥您好,我在几年前曾经首次打过五针法前两针的狂犬疫苗。当时十日观察法医生是告诉我没风险了没有接种。然后前段时间暴露了,去接种,医生和我说我以前打过疫苗,统一接种加强针即可,所以只给我打了两针。
请问一下,我以前只打了五针法的前面两针,没把全程打完。这次再暴露,像医生说的接种两针加强针就够了吗?我要不要再去接种三针啊。
你遇到了一个非常好的医生,我也赞同这么做