
Contents
The Challenge of Lyssavirus Infections in Domestic and Other Animals: A Mix of Virological Confusion, Consternation, Chagrin, and Curiosity
Rupprecht, C.E.; Belsare, A.V.; Cliquet, F.; Mshelbwala, P.P.; Seetahal, J.F.R.; Wicker, V.V. The Challenge of Lyssavirus Infections in Domestic and Other Animals: A Mix of Virological Confusion, Consternation, Chagrin, and Curiosity. Pathogens 2025, 14, 586. https://doi.org/10.3390/pathogens14060586
1. 引言
关于传染病的经常重复引用的起源仍然是推测性的(即,“……现在是合上传染病之书并宣布抗击瘟疫之战胜利的时候了……“),但总的来说,”微生物战争“才刚刚开始。在人类医学和兽医学中,传染病的“清单”对应届毕业生和经验丰富的专业人士构成了挑战。这部分是由于进化、气候变化、环境退化、全球化、技术、信息流行病等。
从广义上讲,此类病原体(即来自希腊语“痛苦生产者”)的流行病学范围多种多样:从认识来看,经过数千年到新出现;发生率从频繁到罕见;在分销方面,从世界各地到地方;在系统发育宿主宽度上,从宽到窄;发病率从轻度到重度;在生物医学反应方面,从容易识别和治疗到诊断困境和无法治愈;在政治支持方面,从优先次序到被忽视;用学术术语来说,从普通的教学到几乎晦涩难懂的;具有全球意义,从酝酿的大流行潜力到迅速被遗忘、灭绝的现象学;以及介于两者之间的所有内容。跨学科、同一个健康方面,特别是关于对个体生物安全的威胁、种间传播性、食品安全、健康差异、兽医经济学、人畜共患风险、生物政治学和其他主观标准,有助于证明将主题纳入学生基础课程空间的合理性,并扩大了对持续专业培训的需求。
考虑到这种混合,狂犬病病毒是在有点复杂和具有挑战性的微生物学领域中讨论的理想病毒病原体。狂犬病病毒是同一疾病的唯一病原体,历史上称为狂犬病(即狂暴)。从本质上讲,这些病原体分布于全球,其中不同的哺乳动物宿主对人类、家畜和野生动物构成病毒暴露的威胁。此外,尽管这种疾病已经被识别了数千年,但它在分类学水平上的离散和可变的因果关系是在不到50 年前被发现的,这造成了相当大的困惑、惊愕、懊恼和好奇。
我们综述的目的是从科学的角度看待全球范围内狂犬病病毒在现代兽医学中面临的挑战,通过选定的案例进行说明,并得到同行评审文献中关键历史引文和相关文章的支持。
2.病原体和分类学
兽医学对疾病(例如狂犬病)和病因学(例如丽莎病毒)的潜在混淆部分在于对应用于病毒学领域的现代分类法(即来自希腊语的“顺序”或安排“和”方法“或”规律“)的理解。虽然哺乳动物等生物体(例如,家养食肉动物、狗、犬科、家犬)由于共同特征(理想情况下反映了进化关系)而被分类和排列,但病毒被分为不同的组,基本上基于结构和生化特性,使用二项式命名法的做法。病毒分类学的总体意图是在该领域实现更大的一致性、识别度、沟通性和清晰度 [19]。Lyssaviruses 是杆状、单链、负链 RNA 病毒。在分类学上,它们代表了一个独特的单系类群,居住在弹状病毒科(Lyssavirus)属,其划分标准部分基于核苷酸身份、血清学和生态位空间[20]。所有丽莎病毒病毒粒子都具有相似的子弹状形态(~60-110 nm × ~130-250 nm)。核酸包含在内部螺旋状核衣壳中,具有外部膜结合蛋白外壳(图 1)。

图 1.广义子弹状丽莎病毒病毒粒子的概念,说明了外部 G 蛋白尖峰(紫色),延伸穿过宿主细胞衍生的膜(白色),带有 M 蛋白(黑色和红色珠状点画)的剖面图,包裹着螺旋核糖核蛋白核心,由包裹在单链 RNA 上的 N 蛋白(灰色)组成,P 蛋白(品红色), 和 L 蛋白 (teal),也称为 RNA 依赖性 RNA 聚合酶(由 I. Kuzmin 提供,德克萨斯大学医学分部,加尔维斯顿,德克萨斯州,美国)。
五个单顺反子基因编码结构蛋白,包括一个核蛋白 (N);磷蛋白 (P);基质蛋白 (M);一种糖蛋白 (G);和“大”RNA 依赖性 RNA 聚合酶 (L)。转录和复制都发生在宿主细胞细胞质中(通常但不限于神经元)。病毒多肽在病毒周期的基本结构和关键方面具有多种作用,从最初的细胞接收到转录和翻译,再到病毒粒子组装和病毒排出。通过长期适应脊椎动物宿主,丽莎病毒进化出双重隐身和抑制机制,以保留神经通路、抑制细胞凋亡并限制宿主炎症反应,包括调节干扰素通路。正在进行的全球研究为这些病原体在体内利用的优雅机制提供了关键见解,这些机制在哺乳动物 CNS 的基本生态位中随时间和时间延续。
| Lyssavirus (系统群) | 常用名 | 宿主 | 年份和 地区 | 溢出 感染 |
|---|---|---|---|---|
| Lyssavirus aravan (I) | Aravan 病毒 (ARAV) | 食虫蝙蝠(例如 Myotis blythi) | 1991 年,中亚(例如,吉尔吉斯斯坦南部) | ? |
| Lyssavirus australis (I) | 澳大利亚蝙蝠狂犬病病毒 (ABLV) | 食虫和食虫蝙蝠(例如 Pteropus、Saccolaimus 等) | 1995 年,澳大利亚 | 人类、马 |
| Lyssavirus bokeloh (I) | Bokeloh 蝙蝠狂犬病病毒 (BBLV) | 食虫蝙蝠(例如 Myotis、Pipistrellus spp.) | 2009 年,欧洲(例如,德国、法国、波兰等) | ? |
| Lyssavirus caucasicus ((III)) | 西高加索蝙蝠病毒 (WCBV) | 食虫蝙蝠(例如 Miniopterus schreibersii) | 2002 年,欧洲(例如,俄罗斯克拉斯诺达尔地区、意大利等) | 猫 |
| 杜文哈格 莱萨病毒 (I) | Duvenhage 病毒 (DUVV) | 非洲蝙蝠(例如,Nycteris thebaica) | 1970 年,撒哈拉以南非洲(例如,南非、肯尼亚、津巴布韦等) | 人类 |
| Lyssavirus formosa (I) | 台湾蝙蝠狂犬病病毒 (TWBLV) | 食虫蝙蝠(例如 Pipistrellus spp.) | 2016, 中国台湾 | ? |
| Lyssavirus gannoruwa (I) | Gannoruwa 蝙蝠 lyssavirus (GBLV) | 果蝠(例如,Pteropus spp.) | 2014, 斯里兰卡 | ? |
| Lyssavirus hamburg (I) | 欧洲蝙蝠狂犬病病毒 1 型 (EBLV1) | 食虫蝙蝠(例如 Eptesicus、Myotis、Rhinolophus spp. 等) | 1950 年代,欧洲(例如比利时、丹麦、法国、德国、荷兰、波兰、俄罗斯、西班牙、乌克兰等) | 人类, 猫, 羊, 貂 |
| Lyssavirus 赫尔辛基 (I) | 欧洲蝙蝠狂犬病病毒 2 型 (EBLV2) | 食虫蝙蝠(例如 Myotis spp. 等) | 1985 年,欧洲(例如,芬兰、荷兰、瑞士、英国等) | 人类 |
| Lyssavirus ikoma (III) | 生驹狂犬病病毒 (IKOV) | 未知 | 2009 年,坦桑尼亚 | 灵猫 |
| Lyssavirus irkut (I) | 伊尔库特病毒 (IRKV) | 食虫蝙蝠(例如 Murina leucogaster) | 2002 年,欧亚大陆(例如,伊尔库茨克、俄罗斯东西伯利亚地区、中国) | 人类, 狗 |
| Lyssavirus khujand (I) | 苦盏病毒 (KHUV) | 食虫蝙蝠(例如 Myotis spp.) | 2001 年,中亚(例如,塔吉克斯坦北部) | ? |
| Lyssavirus Kotalahti (I) | Kotalahti bat lyssavirus (KBLV) | 食虫蝙蝠(例如 Myotis spp.) | 2017, 芬兰 | ? |
| Lyssavirus lagos (II) | 拉各斯蝙蝠病毒 (LBV) | 非洲蝙蝠(例如,Eidolon helvum、Epomophorus wahlbergi、Micropteropus pussilus、Rousettus aegyptiacus 等) | 1956 年,撒哈拉以南非洲(例如,中非共和国、埃塞俄比亚、加纳、几内亚、肯尼亚、尼日利亚、塞内加尔、南非、津巴布韦等) | 猫、狗、水猫鼬 |
| Lyssavirus lleida (III) | 莱里达蝙蝠狂犬病病毒 (LLEBV) | 食虫蝙蝠(例如,Miniopterus spp. | 欧洲(例如西班牙、法国等) | ? |
| Lyssavirus mokola(II) | Mokola 病毒 (MOK) | 未知,但怀疑是小型哺乳动物 | 1968 年,撒哈拉以南非洲(如喀麦隆、中非共和国、埃塞俄比亚、尼日利亚、南非、津巴布韦等) | 猫、狗、人类等。 |
| Lyssavirus rabies (I) | 狂犬病病毒 (RABV) | 蝙蝠、中食肉动物、非人灵长类动物 | 据信,在人类历史上,数千年来一直得到认可 | 理论上,所有温血脊椎动物 |
| Lyssavirus shimoni (II) | 希莫尼蝙蝠病毒 (SHIBV) | 非洲蝙蝠(例如,Macronycteris vittatus) | 2009 年,肯尼亚 | ? |
| UNCL | Divača bat lyssavirus (DBLV) | 食虫蝙蝠(例如 Myotis capaccinii) | 2014, 斯洛文尼亚 | ? |
| UNCL | Phala bat lyssavirus (PBLV) | 蝙蝠(例如 Nycticeinops schlieffeni) | 2021, 南非 | ? |
| UNCL | 台湾蝙蝠狂犬病病毒 2 型 (TWBLV2) | 蝙蝠 (Nyctalus plancyi velutinus) | 2018, 中国台湾 | ? |
| UNCL | Matlo bat lyssavirus (MBLV) | 非洲蝙蝠(例如,Miniopterus natalensis) | 2015, 南非 | ? |
3. “狂犬病相关病毒”的历史
几千年来,多种文化都感觉到动物咬伤与随后导致死亡的疾病之间存在相关性。“狂犬病”(即在动物中“愤怒”或施暴)或“恐水症”是这种可识别现象的通用名称,即使并非所有人都同意最终的因果关系。这种“可传播的毒素”甚至可能是“病毒”一词通用定义的基本原理,广义上解释为“粘稠液体”或“毒药”。几个世纪以来,随着科学理念的不断发展以及显微镜和实验室方法技术的逐渐进步,看似较大的病原体,如细菌和真菌,变得可以与“病毒”区分开来,即所谓的“无限小”的不可过滤微生物(或到 19 世纪末,Beijerinck 经典地认为是“contagium vivum fluidum”)。到 20 世纪中叶,通过电子显微镜观察烟草花叶病(Tobamovirus tabaci)、口蹄疫(口蹄疫病毒)、狂犬病(Lyssavirus rabies)和许多其他病原体的颗粒性质。命运、天体事件、瘴气、自发产生和无数其他解释都被蓬勃发展的病毒学所取代。
最初,人们认为只有狂犬病病毒 (RABV) 才会引起狂犬病。然而,这种想法在 20 世纪中叶发生了变化,当时蝙蝠被认为是潜在的病毒宿主(图 2a、b)。现场和实验室方法的混合打破了 RABV 的垄断地位,人们认识到密切相关的病毒可能导致相同的疾病。
图 2.(a) 在 20 世纪,在考虑病原体延续领域的潜在病毒库时,理想的宿主特征,如高敏捷性、极端生物多样性、广泛分布、社会群居性以及蝙蝠固有的其他几个属性,越来越受到重视。(b) 一只蝙蝠在房屋阁楼上休眠时的特写照片(由 ME 提供。Rupprecht, Dacula, GA, USA)。
尽管来自新世界的历史记载表明,自欧洲殖民时期以来,吸血蝙蝠 (Desmodus rotundus) 就参与了吸血蝙蝠的参与,但关于手翅目在病毒性人畜共患病或新发传染病 (EID) 中的任何作用的记录等待改进的诊断方法的出现。在 1950 年代,实地调查开始在不同动物中寻找与脑炎有关的各种因子。这些全球研究涉及组织收集和病毒分离尝试。其中一次“病毒狩猎”探险发生在尼日利亚的拉各斯岛,包括对果蝠的投机取样。这些蝙蝠的脑匀浆 (Eidolon helvum) 在小鼠脑内接种后是致命的。这种分离的试剂称为拉各斯蝙蝠病毒 (LBV),通过电子显微镜呈子弹状,类似于 RABV 病毒粒子(图 3)。
图 3.透射电子显微镜图像显示了 LBV 病毒粒子的出芽子弹状颗粒,外围围绕着中央卵胞浆内包涵体(由 F. Murphy、S. Whitfield、美国 HHS 公共卫生图像库提供)。
在1960年代,从尼日利亚鼩鼱中获得的另一种分离株也杀死了实验室小鼠。这种分离株的名称是 Mokola 病毒 (MOK),一种疑似人畜共患病原体。LBV 和 MOK 在形态学上与 RABV 相似,但可通过血清学区分,分别称为狂犬病血清型 2 和 3 。在实验性感染期间,接种 RABV 疫苗的小鼠对 MOK 的抵抗程度最低 。LBV 和 MOK 在接种到狗和非人灵长类动物中时都会产生致命的感染。比较 RABV、LBV 和 MOK,病毒核糖核蛋白 (RNP) 表现为统一的“狂犬病组特异性抗原”,而包膜相关抗原(G 蛋白)被认为是成员中的血清型决定因素。
lyssaviruses 的名单随着额外的流行病学内省而增加。在南非,一例“人类狂犬病”发生在一名被蝙蝠咬伤的男子嘴唇上,但免疫荧光反应性似乎不典型。Duvenhage 病毒 (DUVV) 是分配给该分离株的名称,以死于狂犬病的患者的名字命名。考虑到病毒粒子形态、对超免疫血清的反应原性以及相关的脑炎感染,DUVV 加入了“狂犬病血清组”,作为血清型 4。
1978年,美国宾夕法尼亚州费城Wistar研究所的研究人员报道了产生针对RABV和相关病毒的单克隆抗体(MAb)的杂交瘤。这些 MAb 识别 RABV 的抗原变体,并将其与 DUVV、LBV 和 MOK 区分开来。针对病毒 N 和 G 蛋白的 MAb 面板允许对人类、家畜和野生动物的分离株进行抗原分型。虽然病因学上的组内差异很明显,但从不同宿主的时空获得的不同病毒自然或实验上导致了相同的致命疾病。即病毒接种后,实验动物发展为急性进行性神经系统疾病,脑实质、胞质内包涵体中轻中度炎性浸润,电镜观察观察到典型的弹状病毒颗粒在神经元内质网和质膜上出芽。
从1968年到1985年,欧洲蝙蝠和芬兰的一位蝙蝠生物学家又发现了病毒分离株,后者在被蝙蝠(Myotis daubentoni)咬伤7周后出现神经系统症状,23天后死亡。最初,根据 MAb 分型,这些分离株被认为是与非洲 DUVV 相关的地理欧洲亚群(提出血清型 5)。三十多年来,“狂犬病相关病毒”一词起源于应用研究,基于临床疾病、病理表现、形态学表现、血清学交叉中和和抗原分化。渐渐地,人们意识到狂犬病不仅仅是由 RABV 引起的。从 1980 年代至今,后来使用现代系统发育方法所获得的明显好处进一步证明了狂犬病是由于全球不同哺乳动物种群中长期存在的不同狂犬病病毒感染所致(表 1)。
目前,病毒分类学家识别出 >20 种推定的狂犬病病毒物种。随着病原体发现和科学好奇心的进行,预计会有更多人。在 lyssaviruses 中,基于遗传差异的物种划分标准被认为比其他一些病毒属短一些。从本质上讲,这意味着在某个时候,所有 lyssaviruses 都可能被简单地称为 RABV。仅根据系统发育原因进行分裂,从来不意味着这些病毒物种在它们在各自宿主中产生的疾病或构成人畜共患病的公共卫生问题上有所不同。某些生物医学专业人士的困惑是可以理解的,他们可能没有意识到这个基本点。然而,当今文献中普遍存在误解,当人们读到“狂犬病是由 RABV 引起的”时,尽管自 20 世纪中叶以来的数据相互矛盾。这种分类多样性在 RNA 病毒中很常见,并且在缺乏校对酶的情况下是意料之中的:一些突变导致病毒灭绝;许多变化是中性的;其他疾病则为适应和疾病出现提供了选择性机会。这是介绍分配给 lyssaviruses 的应用命名法历史的关键,在更大的疾病认识领域,适用于兽医学、公共卫生和保护生物学。
正如这位吟游诗人曾经反问的那样,“……名字里有什么……?在讨论未来的疾病命名法时,显然应该避免使用患者名字(例如,“Duvenhage”)。同样,延续“地名”(例如,“拉各斯蝙蝠病毒”)的好处微乎其微。这种使用会造成污名化、偏见和混淆。此外,随着广泛的全球化、气候变化、宿主跳跃、病原体易位等的实现,随着疾病的扩大(或意识到比人们认为的要广泛得多),这种狭窄的地理分配变得毫无意义。使用像“Newville Pitbull 圣诞蝙蝠病毒病”这样的奇特术语来描述最初在一个小镇范围内报告的虚构疾病,并在 3 个月前食用了一只蝙蝠后从单一品种的小狗中季节性分离出来,当一种新的人畜共患病后来在整个地区爆发时,变得有限、多余和具有误导性, 影响狗、猫、家畜和家禽(实际上与蝙蝠无关)。为避免错误信息,理想的替代方案应简单、准确、具有交流性,并且与明显的临床术语更相关。
微生物领域固有的,不同的寄生虫、真菌、原生动物、细菌和病毒类群与它们各自的描述性疾病有关(补充表 S1)。因此,“狂犬病病毒”(即病因)和“狂犬病”(即疾病)之间的关系没有什么不同——狂犬病是一种由 Lyssavirus 属 RNA 病毒引起的病毒性疾病。因此,那名因蝙蝠咬伤而得名的南非男子并没有死于“非洲杜文哈奇蝙蝠病毒”,而是死于狂犬病(正如Merdith等人在1971年的原始病例报告标题中所暗示的那样)。
4. 宿主/病原体和临床谱
所有狂犬病病毒都会引起急性进行性脑炎。无论病毒种类如何,临床体征都高度可变,但基本上无法区分。所有温血脊椎动物都可以支持生产性病毒感染。鸟类易感,但报告很少见,最近没有进行比较调查。在哺乳动物中,负责种内延续和种间溢出感染的野生动物宿主存在于手翅目、食肉动物和灵长类动物中。其他哺乳动物物种则是媒介或受害者。在家养哺乳动物中,狗是独一无二的,它是狂犬病病毒宿主、媒介和受害者的唯一代表。猫科动物是有效的媒介,但不是宿主,并且在该病的流行病学中越来越重要。今天,在低收入和中等收入国家 (LMIC) 无人监督的社区犬构成了与狂犬病相关的最重要的公共卫生问题。
奇怪的是,感染报告涵盖了家养物种的广泛范围,从伴侣动物到牲畜,以及圈养的外来分类群,从小型袖珍宠物到大象(补充表 S2)。这种多样性影响了传统从业者、动物园兽医和专门研究管理野生动物健康的人。如果迄今为止哺乳动物物种的相关病例报告很少(例如有袋动物)或缺乏(例如鲸类),这可能表明存在潜在的生物学/生态学因素或反映缺乏流行病学内省。在已描述的类群中,狂犬病的临床谱范围较广:从前驱期的非特异性表现,到急性神经期的显著症状(即所谓的 “狂躁型或麻痹型” 表现),随后会出现昏迷并最终导致死亡。这一临床过程的差异,部分取决于病毒剂量、感染途径、病情严重程度、宿主物种,以及专业人员通过敏锐观察所具备的鉴别能力(补充表 S3)。
可以理解的是,与 lyssavirus 感染相关的临床症状差异很大。虽然从疯狗身上召唤出的躁狂症形象激发了媒体和公众的想象力,但发作是非特异性的,麻痹的形式要微妙得多。此外,畜牧业的病例经常被忽视。然而,一些迹象,例如发声改变,如牛不停地咆哮,几乎是特异性的,一旦听到,就很难忘记。此外,体型较大的动物的脑炎是一种危险的组合。人们被狂犬病牛和马严重伤害或直接杀死。
尽管狂犬病病毒是与牲畜狂犬病病例最常见的病原体(尤其是在拉丁美洲,由于受感染的吸血蝙蝠的捕食),但在家畜中也发现了其他狂犬病病毒。例如,欧洲猫中报道了 EBLV 和 WCBV。在接种疫苗的非洲猫和狗中观察到 MOK 和 LBV。在欧洲,1998 年至 2002 年间,在 5 只患有神经系统疾病的丹麦绵羊中发现了 EBLV1。在三只患病动物起源的牛群中的两个牛群中,在 69 只绵羊中的一只中检测到 EBLV1 中和抗体。2000 年,在屠宰场收集的 >2000 份绵羊血清的 EBLV1 抗体均呈阴性,在 87 只表现出神经系统体征的动物中未发现 EBLV1。虽然除RABV外的其他狂犬病病毒在自然条件下确实会感染牲畜,如绵羊,但在欧洲,此类事件仅作为偶然事件出现。大多数狂犬病病毒在种内传播,具有不可预测的溢出感染机会,可对人类、家畜和野生动物造成溢出感染(图 4)。单一病毒种属分离且间隔时间长而没有额外报告意味着监测不足或病毒灭绝的可能性。
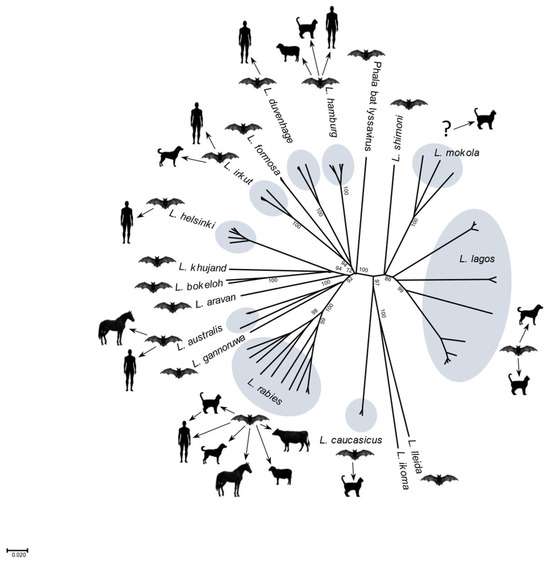
图 4.所选 lyssavirus 物种的系统图,基于已知或假定的哺乳动物宿主和溢出感染到其他物种的迹象(由美国德克萨斯州加尔维斯顿德克萨斯大学医学分部的 L. Kuzmin 提供)。
5. 狂犬病病理生物学
简而言之,狂犬病病毒进入宿主外周组织,在 CNS 中复制,并通过唾液排泄。病毒暴露定义为直接经皮或黏膜接触已知或可能感染的物质,如唾液、唾液腺、脑或相关神经组织。尽管有多种潜在的非咬伤暴露途径,但几乎所有狂犬病病例都是由咬伤引起的。患有狂犬病的动物会分泌大量的病毒粒子并产生大量的唾液。非咬伤暴露包括抓伤、吸入气溶胶、开放性病灶污染或粘膜接触传染性物质,例如从唾液中排泄病毒的狂犬哺乳动物舔到新鲜伤口,或直接接触鼻子、嘴巴、眼睛等。暴露后,潜伏期从几天到几个月不等(在极少数情况下> 1 年不等)。长度部分取决于病毒剂量和途径。描述了无数的临床体征,没有一个是具有特异性的,超出了规范中的非特征行为症状(补充表 S3,补充图 S1)。在某些情况下,急性死亡可能没有观察到任何体征,而在其他情况下,3-4 天的发病期可能表现为愤怒或瘫痪表现 。根据自然感染和实验研究,在狗、猫和雪貂中,根据自然感染和实验研究,病毒排泄可能不会发生,可能与临床症状同时出现,也可能在发病前7至10天排出。缺乏来自其他物种的比较数据。虽然一些狂犬病病例可能看起来相当戏剧性,但其他病例更为微妙,仅靠观察不足以确认,因为疑似脑炎的鉴别诊断范围很广,包括其他相关的感染性病因、毒性和创伤。
在极少数情况下,狂犬病病毒可在 CNS 生产性感染期间诱导主要结构变化。然而,与高病死率和通常明显的临床体征相比,显微镜检查时的病理变化通常很小。当狂犬病不在鉴别诊断清单上时,兽医病理学家可能很容易漏诊病例。大体病变不明显,但可能包括自我创伤、吸入性肺炎、脑膜充血或脊髓出血的证据。从历史上看,在 CNS 内,微观血管周围浸润或“Babes 结节”,以及 Van Gehutchten 对退行性神经节细胞变化的描述,在某些情况下被注意到,但被认为是非特异性的,也不是普遍的(补充图 S2)。20世纪初,意大利病理学家Adelchi Negri在狂犬病动物的脑涂片中检测到椭圆形或球形包涵体,认为它们表明寄生微生物,如原生动物。与此同时,美国病理学家Anna Wessels Williams也观察到了类似的显微镜观察结果。然而,内格里首先公布了他的结果。因此,这些结构以他的名字命名。这些胞浆内包涵体或“Negri 小体”(而不是“Williams 小体”)可能在受感染的神经元中发现,大小和数量波动,或者可能不会发生(补充图 S3)。
尽管它们的生物地理和抗原/遗传多样性不同,但与狂犬病病毒相关的临床病理光谱难以区分,无论是在其自然宿主中还是在实验性感染期间(例如,RABV、EBLV、DUVV、WCBV),如使用免疫组织化学 (IHC) 或类似技术观察到的那样(图 5a-e)。通过比较狂犬病病毒种类、分离来源、暴露途径、病毒剂量和动物模型,可以在实验中观察到各种体征、潜伏期、脱落、存活、免疫反应等,这表明不同的致病指数,外推到自然宿主。
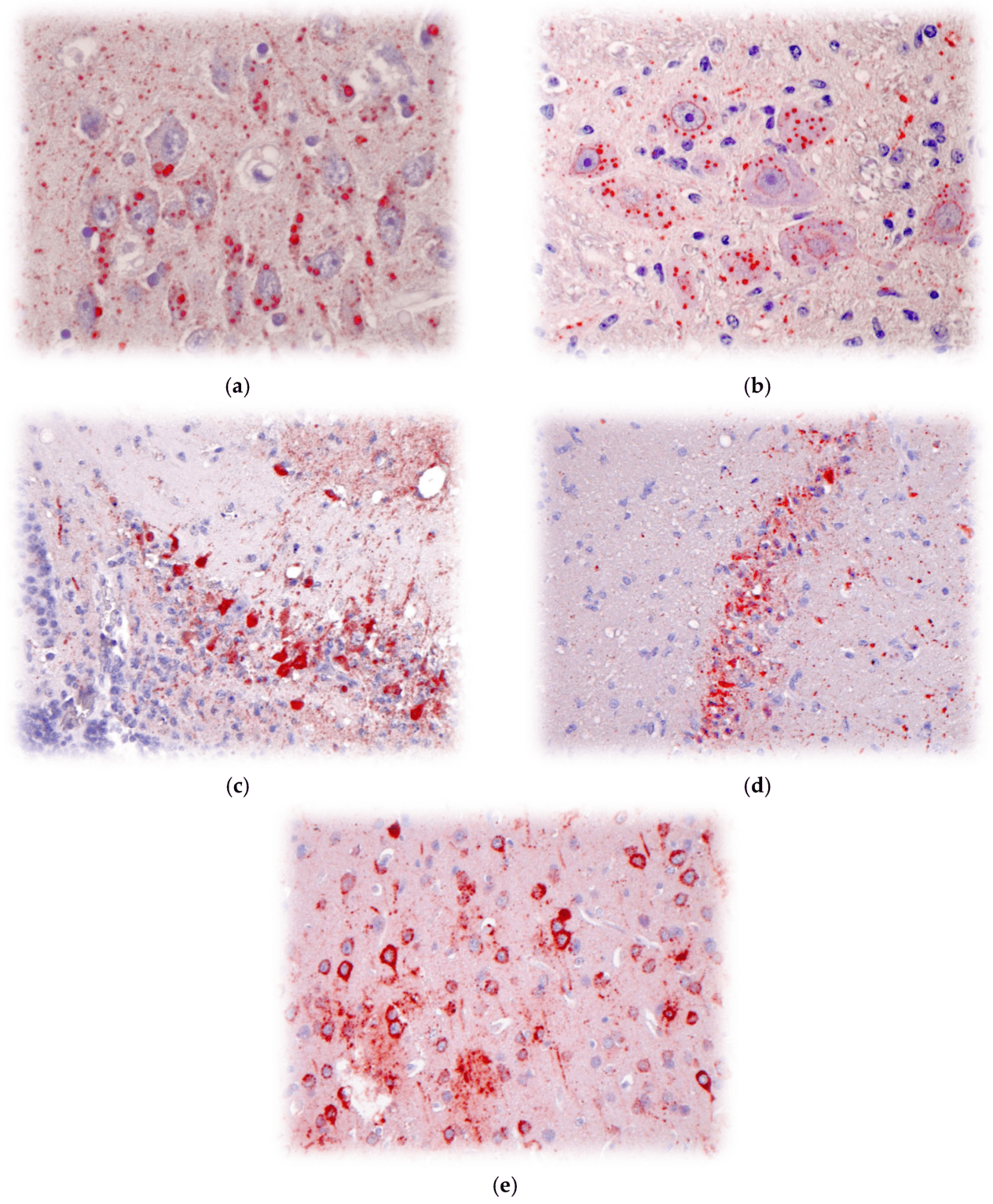
图 5.(a) IHC 在显微镜下检测到患有狂犬病的狗的脑组织中丰富的 RABV 抗原(洋红色染色的包涵体),400×(由美国佐治亚州亚特兰大美国疾病预防控制中心的 M. Niezgoda 提供)。(b) 通过 IHC 在显微镜下检测到狂犬病羊脑组织中丰富的 RABV 抗原(品红色染色包涵体),400×(由 M. Niezgoda,美国疾病预防控制中心提供,美国佐治亚州亚特兰大)。(c) 实验性啮齿动物感染后,IHC 检测到脑组织中丰富的 EBLV1 抗原(洋红色染色包涵体),200×(由美国佐治亚州亚特兰大美国疾病预防控制中心的 M. Niezgoda 提供)。(d) 实验性啮齿动物感染后,IHC 在显微镜下检测到脑组织中丰富的 DUVV 抗原(洋红色染色的包涵体),200×(由美国佐治亚州亚特兰大美国疾病预防控制中心的 M. Niezgoda 提供)。(e) 实验性啮齿动物感染后,IHC 在显微镜下检测到脑组织中丰富的 WCBV 抗原(洋红色染色包涵体),200×(由美国佐治亚州亚特兰大美国疾病预防控制中心的 M. Niezgoda 提供)。
除了单核血管周围袖套、神经胶质结节(局限性神经元变性)、神经吞噬和坏死的组织学证据外,家畜中枢神经系统的其他病理变化可能包括灰质神经细胞的海绵状变化。由于在神经元功能障碍期间组织病理学病变可能不存在、轻微或遗漏,因此需要特异性诊断性检查以确认临床怀疑 lyssavirus 感染。
6. 动物狂犬病的实验室诊断
对狂犬病病毒的可靠检测依赖于能够识别和确认狂犬病病例的诊断检测网络。通常,通过两条主要途径检测动物是否存在 狂犬病 感染:被确定为对人类或动物健康构成威胁的对象(通常通过被动监测);以及野生动物监测(通常通过加强监测)。对于具有相容体征的异常嫌疑人,已知在过去几周至数月内接触过经实验室确诊的患有狂犬病的动物(例如,一只现在消瘦、瘫痪、未接种疫苗的户外猫,在捕捉到检测呈阳性的病蝙蝠 6 周后逃脱但再次出现在“主人”的场所),诊断相当简单。不幸的是,事情很少简单,因为前驱症状的发作是非特异性的,而且许多暴露可能未被识别,尤其是在自由放养的动物中。在人类中,在国家参考中心进行的生前方法可用于疑似病毒性脑炎患者。在其他动物中,出于各种原因,尸检技术是常态。值得注意的是,患有狂犬病的动物对主人和兽医人员构成重大的公共卫生危害。考虑到医疗选择、攻击性行为、自残、终末期痛苦等,为兽医安乐死提供了合乎逻辑的理由,因为发病期可能很短且极端,认识到这种疾病是无法治疗的。重要的是,如果动物暴露了人,作为人类暴露后预防(PEP)风险评估的一部分,包括伤口护理、疫苗接种以及针对未接受过治疗的患者的咬伤狂犬病免疫球蛋白(RIG)/MAbs。此外,当地动物卫生法规可能排除某些类型的体内检测的应用(评估对疫苗接种的免疫反应或血清学监测以获得流行动物学见解除外)。
在过去的75年里,各种实验室技术不断发展起来,用于动物狂犬病的适当尸检诊断,重点是检测裂解病毒抗原、核酸和抗体。此类测试灵敏、特异、快速且经济,取决于适合目的的需要 。此外,这些测试可以作为“筛选”相互结合使用,以降低总体检测成本。如果组织学提示狂犬病病毒感染,特异性实验室检查可以确认最初的怀疑。如果新鲜/冷冻样本不可用,病理组织块可通过 IHC 提供机会(图 5)。其他经典方法,如电子显微镜(图3)、病毒分离等,可能对应用研究有用,但目前很少用于常规诊断应用。
实验室检测狂犬病的主要考虑因素应该是为检测而采集的样本类型。由于狂犬病病毒具有嗜神经性,且病毒在中枢神经系统复制后开始发病,因此脑干的完整横截面(以及小脑的所有三个叶)是最可靠的诊断样本,是明确排除感染所必需的。CNS 的其他部分可能为阴性,而脑干检测为阳性。理论上可以对不同组织类型(例如皮肤活检、唾液、脑脊液、血清等)进行生前检测,但这种方法通常仅用于人类疑似狂犬病病例。生前检测所需的组织对患者具有侵入性,由于难以及时准确采集和检测这些材料,以及非神经元组织中可能缺乏分析物,因此可能无法排除狂犬病。在所有情况下,正确采集、储存和快速运输样品对于保持不同检测方法所针对的分析物的完整性都至关重要。样品到达实验室后,必须小心处理样品,并消除所有交叉污染源。
诊断和监测试验的结果可以确认 狂犬病 感染,有时可以排除 狂犬病 感染,或导致不确定的结果。不确定的结果和低置信度的阴性结果(例如,侧向层析试验的阴性结果,LFA),应送至参考实验室进行确认性检测。这种二次测试可能会解决或维持不确定的结果,具体取决于样品的条件和类型(图 6)。
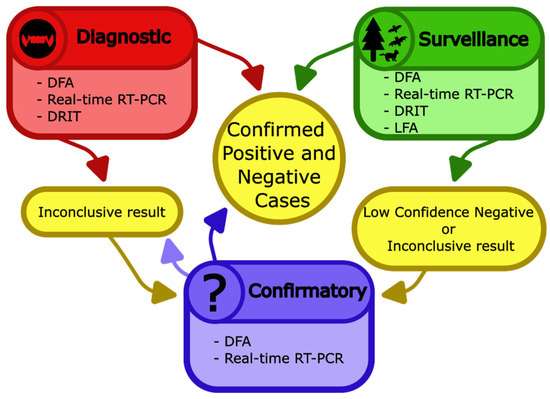
图 6.用于检测动物 狂犬病 的实验室和现场测试。
6.1. 基于核酸的检测
基于核酸的检测旨在提取和扩增样品中的靶 RNA。基于核酸的检测的一个主要优点是可以灵活地特异性靶向 狂犬病 属的广泛和狭窄代表。实验室开发的检测范围从检测RABV的亚变体到检测所有已知的狂犬病种属。
常规 RT-PCR 是这些类型测试中的第一种。简而言之,提取的 RNA 被扩增以制造 cDNA,然后通过电泳在凝胶中观察 。后来,这种方法发展为半巢式RT-PCR,其中来自RT-PCR的cDNA扩增子被用作额外PCR反应的模板DNA。半嵌套方法更敏感,几十年来一直用于确认世界各地的狂犬病病例。然而,它也更容易出现假阳性结果,因为污染物 cDNA 可以在二级反应中被扩增。
最近,PCR 技术的进步导致了两种实时 RT-PCR 泛狂犬病病毒诊断检测方法的发展和改进:JW12 和 LN34。这些检测通过添加荧光染料标记的探针来提高传统 RT-PCR 检测的灵敏度,这些探针在靶 RNA 复制时会发出荧光。此外,实时荧光定量 RT-PCR 可以在单孔、一步反应中完成,从而降低了交叉污染的可能性。这两种检测方法均已经过全面验证,可用于人类生前和动物死后诊断检测,并可作为筛选样本或确认其他诊断检测结果的工具(例如,直接荧光抗体检测,DFA;直接快速免疫组织化学检测,DRIT;等)。
基于核酸的分析的其他进步催生了等温扩增方法,这种方法在资源有限的环境或现场可能更容易获得,因为它们不需要热循环仪来完成扩增反应。已经开发了环介导等温扩增 (LAMP) 和重组酶聚合酶扩增 (RPA) 检测方法来检测 RABV。然而,LAMP和RPA检测通常不能检测所有RABV变异,也没有开发出用于检测其他狂犬病病毒的分析方法。在使用这些方法进行诊断或监测之前,应验证新的等温扩增测定,以确保对所有局部狂犬病病毒的敏感性。
作为一种诊断工具,基于核酸的检测在整个狂犬病病毒领域都有效,尤其是在其他分析物降解或受限的情况下。这些检测方法也被用于检测和表征新型狂犬病病毒,这些病毒在其他诊断性检测中可能无法预测反应。最后,基于核酸的检测可作为其他方法的验证性检测,以解决不确定的结果。在比较研究中,成熟的 RT-PCR 检测显示出与 DFA 相似或更高的敏感性和特异性。与所有诊断检测一样,应定期重新评估基于核酸的检测,以确保正确覆盖目标物种和病毒变体。此外,这些分析的进一步改进通过简化方案和降低成本,使其更高效,更容易为实验室所用。
6.2. 基于抗原的检测
已经开发了基于抗原的检测方法,用于检测和观察组织样本中的狂犬病病毒抗原。这组检测包括 DFA,由于其高特异性和敏感性,它是历史最悠久的狂犬病诊断检测之一。抗原检测的核心方法是采集具有代表性的脑组织样本,应用与 狂犬病 抗原特异性结合的抗体,然后可视化抗原-抗体复合物(图 7)。抗原检测依赖于可用组织的质量、合适的抗体的可用性以及进行检测的技术人员的经验。研究表明,分解会降低抗原检测的反应性,从而导致假阴性或不确定的结果。
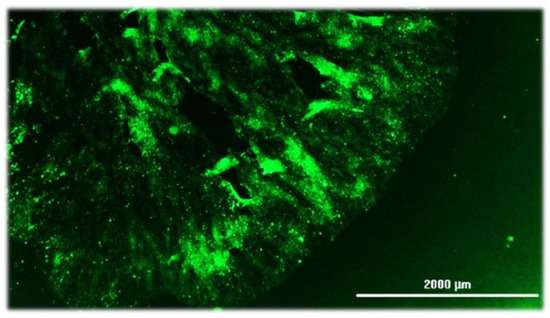
图 7.对自然感染的狂犬病猫的脑印进行直接荧光抗体检测的阳性结果(由美国堪萨斯州曼哈顿堪萨斯州堪萨斯州兽医诊断实验室的 R. Davis 提供)。
在所有基于抗原的检测中,所选抗体必须对所有当地流行的狂犬病病毒具有反应性。因此,实验室必须验证任何新的抗体偶联物,以确保对当地狂犬病病毒种类和变体具有足够的敏感性和特异性。通常,抗原检测中至少使用两种 MAb 偶联物,以确保覆盖局部变异,因为任何单一的 MAb 可能不会对所有已知的 狂犬病 和狂犬病变体产生反应。从历史上看,与 MAb 相比,多克隆抗体对 RABV 变体表现出更广泛的反应性。然而,需要进一步的开发和验证,以确保任何分析物对其他病毒种类和变体的广泛反应性。
DRIT 开发于 20 世纪末,遵循与 DFA 类似的方案,但进行了一些更改,使检测更快速、更容易实施。此协议大约需要一个小时才能完成。使用生物素化的抗 N 蛋白 MAb 或多克隆血清,无需使用荧光显微镜。相反,DRIT 期间产生的脑干印模玻片用光学显微镜分析。独立验证表明,DRIT 和 DFA 之间的敏感性和特异性相似,并且能够检测非 RABV 狂犬病病毒。DRIT 通常用于加强野生动物狂犬病监测和诊断狂犬病病例。然而,与任何诊断性检查一样,不确定的结果应通过确认性检查进一步分析。
与 DFA/DRIT 相比,LFA 是一种使用脑干组织样本进行抗原检测的不同方法,近年来引起了国际关注。LFA 设备的低成本使这种方法具有吸引力,尽管人们对这些检测所有狂犬病存在的测试的可靠性存在挥之不去的担忧。2023年,国际组织发布了一份关于这类诊断设备的声明,警告不要将其用作诊断狂犬病病例的主要方法。
对LFAs的评估显示,根据品牌、批次和对制造商说明的依从性,诊断质量差异很大。令研究人员感到震惊的是,生产 LFA 的公司通常不会披露对样品中存在的抗原反应的特异性抗体。如上所述,抗原检测技术依赖于正确选择对所有本地循环的狂犬病病毒反应的抗体。迄今为止,研究表明对各种溶疮病毒的敏感性相互矛盾。除了担心 RABV 以外的 LFA 对其他不同 狂犬病 的适用性外,严格遵守生物安全和个人防护设备 (PPE) 对于处理感染的 CNS 组织的技术人员来说至关重要。目前的 LFA 通常在现场环境中进行,无需常规实验室生物安全设备,例如 BSC。因此,技术人员应使用适当的 PPE 并仔细对 CNS 组织进行匀浆,以避免气溶胶。技术人员还应意识到,暴露前预防(preexposure prophylsis, PrEP)和PEP(如果有)可能无法针对II.型和III.型的狂犬病病毒提供足够的免疫力。
虽然 LFA 不适合排除 狂犬病 感染,但它们已成为一种很有前途的监测工具,可在进行进一步评估之前筛选现场样本。在这方面,许多地区已经实施了LFAs来检测阳性病例,同时将潜在的阴性样本路由到更可靠的诊断方法。由于用于生产这些装置的抗体的局限性,需要更多的研究来确定某些品牌的 LFA 是否可用于筛选除 RABV 之外的狂犬病病毒物种。此外,出于病理生物学和检测验证问题,它们不应用于唾液或其他不合适的样本类型的尝试检测。
6.3. 基于血清的检测
血清学检测通常不用于兽医诊断检测本身,因为通常仅在狂犬病极晚期进展中才存在足够的抗体(如果有的话)。动物通常在目前的诊断方法能够识别出阳性之前就死亡,因为不同物种对各种狂犬病病毒、暴露途径和剂量等的血清转化和个体免疫反应较晚。在人类中,血清学检测用于生前人体检测,以评估 CSF 和血清中的免疫反应。血清学检测(如快速荧光病灶抑制试验 (RFFIT) 或荧光抗体病毒中和试验 (FAVN))与其他检测方法(如实时荧光 RT-PCR 和 DFA)相结合,有助于形成一种强大的检测方法,以评估生前(或死后)样品中的狂犬病病毒活性。
在 FAVN 和 RFFIT 等检测中检测 VNA 使用患者血清或 CSF(和阳性/阴性对照抗体)的稀释液来灭活测试中标准剂量的具有复制能力的狂犬病病毒。将幼稚细胞添加到样品/病毒混合物中,任何剩余的未中和病毒都会在规定的时间内感染和复制。感染的细胞被固定,偶联抗体用于可视化病毒抗原(如 DFA)。对每种稀释度进行评分,以确定与标准阳性和阴性对照相比患者的终点滴度。RFFIT/FAVN 方法可有效评估患者对疫苗接种的免疫反应,有助于生前诊断测试,并且可以在动物种群中提供有用的监测数据。但是,不建议将其作为主要的兽医诊断测试。此外,该方法中使用的抗体偶联物需要与基于抗原的检测类似的考虑。与基于中和的检测相比,很少有ELISA或类似ELISA的检测方法能够检测和诊断狂犬病病毒的多样性。这些类型的检测方法将来可能被证明是抗体检测的更简单的替代方案。然而,在常规使用此类检测之前,需要进一步验证。
随着地理区域朝着控制、预防和选择性消除 RABV 变体(即犬)的目标迈进,加强监测和识别新狂犬病病例的持续能力的需求并未消失。上述诊断方法最适合检测 RABV。许多方法在应用于其他 狂犬病 时结果喜忧参半。这并不奇怪,因为 RABV 是该属中最“成功”的成员,并且在全球范围内对健康的影响最大。然而,随着计划努力实现消除犬类狂犬病的目标,增强检测和预防 狂犬病 传播的能力将至关重要。该领域的进一步研究可能会产生更稳健、更高效的分析方法,用于所有狂犬病的常规检测,并拓宽用于监测目的的可接受样品类型。最后,来自邻近地区和冲突地区的影响在许多地区仍将是一个持续的威胁。因此,在可行的情况下,需要建立强大的诊断网络来阻止狂犬病的传播,并防止病毒在各种生物体和病毒物种之间持续传播。
7. 狂犬病的发生和疾病名称
高度发达国家和低收入国家动物狂犬病的负担往往是未知的。报告可能不需要通报,这在利益相关者、政策制定者和支持者中引起了极大的懊恼。仅基于临床体征的病例严重低估了真实发病率。监测可能更多地集中在城市地区的伴侣宠物上,但较少集中在农村社区的生产动物上。例如,在已消除犬类狂犬病但野生动物狂犬病持续存在的发达国家(如美国),估计发病率可能很小,每年检测的动物中每100,000头动物中不到0.1例。相比之下,在土耳其等同时存在犬类和野生动物狂犬病的国家,牲畜的狂犬病病例每年可能从每100,000头约0.1例到3.9例不等。在埃塞俄比亚,犬狂犬病的估计发病率为每100,000只狗413例,对畜牧业畜牧业生产有很大的经济影响。在该国的另一个地区,2017年至2021年每10万人口中记录了44.9例人类病例和每10万人口3.4例动物病例,这表明公共卫生和农业部门报告之间存在明显且频繁的脱节。
按物种和地区划分,监测偏倚很明显。犬狂犬病的暴发可能影响多种牲畜,如中国,从亚洲农村地区观察到的那样。在犬类和野生动物狂犬病与半游牧文化同时存在的地区,病例普遍存在。与牛相比,猪狂犬病的报告通常不常见,尽管偶尔会发生局部暴发。在森林狂犬病占主导地位的阿曼,山羊和骆驼是 2017-2019 年期间最常报告的狂犬病动物 。自1961年苏丹首次报道狂犬病以来,马类狂犬病的记录一直存在。尽管漏报了,但无论地方性动物病周期涉及患有狂犬病的狗(例如印度次大陆)还是野生动物宿主(例如南美洲),对牲畜的溢出感染并不少见,给当地农业社区造成了重大困难。
在犬类狂犬病已被消除并且通过口服疫苗接种控制野生食肉动物狂犬病的地区,狂犬病总体上可能非常罕见,例如偶尔会通过狂犬病食虫蝙蝠(例如西欧)溢出。一个主要的例外是,在美洲,从墨西哥到阿根廷,牛麻痹性狂犬病病例的暴发每年可能达到数万例,继发于吸血蝙蝠感染。巴西的一项研究估计每年有超过 30,000 例牛感染病例。相比之下,从 1999 年到 2022 年 12 月,全国仅记录了 50,944 例食草动物狂犬病病例。通常,即使是一个病例也就是暴发的一个例子,因为牧场可能远离诊断设施,这使得样本采集和运输变得繁重。根据气候变化的预测,吸血蝙蝠的扩张(以及狂犬病的威胁)可能包括阿根廷、智利和美国的更广泛地区。
除了病原体、宿主和环境的动态领域之外,在病毒学、流行病学和分类学的学术领域之外,还存在每个国家与自我报告疾病定义相关的独立哲学的生物政治现实。实际上,很少有地方是绝对“免费”的,这意味着没有通过充分的、持续的基于实验室的监测系统和疾病报告来检测狂犬病病毒。在一个开明的计划下,考虑到南极洲和整个南大洋其他岛屿地区多样化和丰富的海洋哺乳动物,人们可以质疑南极洲明显没有狂犬病,而随着时间的推移没有任何诊断数据。关于大洋洲也可能提出类似的问题。
显然,消除犬类狂犬病是可实现的重要全球里程碑。各国为了获得这种地位并阻止重新引入而花费巨大。国际卫生组织对“无”各个方面的标准各不相同。例如,世界动物卫生组织为陆生动物的国际贸易设定了标准。他们将狂犬病定义为由 RABV 引起的感染,与动物种类无关。“无狂犬病”国家在过去两年中没有本土感染的 RABV 病例。全球大部分负担是由RABV引起的,据报道,哺乳动物的宿主谱很广,但其他狂犬病病毒是个例外。WHO 在某种程度上相似,但具有不同的健康风险,前提是不存在所有狂犬病病毒、野生动物狂犬病(不包括蝙蝠)或狗。因此,根据所选的分类方案,不同地区可能被认为患有狂犬病,也可能不被认为患有狂犬病(表 2)。矛盾的是,这意味着实验室确认的人类、家畜或野生动物因狂犬病病毒感染而死亡不一定会影响一个地区的狂犬病状况。
表 2.不同国际组织考虑的全球选定地区的狂犬病状况。
| 地区 | WOAH——没有狂犬病? | WOAH—狗没有狂犬病吗? | WHO——无食肉狂犬病? | WHO – 无 狂犬病病毒? |
|---|---|---|---|---|
| 澳大利亚 | 是的 | 是的 | 是的 | 否 |
| 加拿大 | 否 | 是的 | 否 | 否 |
| 中国 | 否 | 否 | 否 | 否 |
| 法国 | 是的 | 是的 | 是的 | 否 |
| 印度 | 否 | 否 | 否 | 否 |
| 日本 | 是的 | 是的 | 是的 | 是的 |
| 尼日利亚 | 否 | 否 | 否 | 否 |
| 乌拉圭 | 否 | 是的 | 是的 | 否 |
| 美国 | 否 | 是的 | 否 | 否 |
8动物狂犬病管理
动物狂犬病的管理范围从个体(如伴侣动物)到大小可变的群体(如畜群健康)和广泛的种群(如野生动物疫苗接种计划)。虽然控制的概念包括诊断、安乐死、行动限制、检疫等要素,但也许最重要的管理方面在于在综合国家计划中通过适当的动物 PrEP 进行预防 。多个变量会影响狂犬病疫苗在动物中的反应。例如,即使在单个物种中,狗接种RABV疫苗的成功概率也可能取决于整体健康状况、使用的疫苗类型、效力、狂犬病疫苗接种次数、品种、接种年龄以及接种疫苗后动物可能暴露的天数。
尽管有许多变量,但狂犬病在正确接种疫苗的动物中相当罕见。可能会发生失败,尽管很少发生,因此无论疫苗接种史如何,对于任何具有与狂犬病相符的临床症状的动物,都应将其纳入鉴别诊断中。必须继续监测以记录此类病例并确定现场规划中的流行病学趋势。即使针对同源病毒,疫苗也可能失败,但可能对异源狂犬病病毒更可能失败,如现场LBV和MOK以及实验室中的其他病毒。开发泛病毒疫苗的尝试仍在继续。
在过去的一个世纪里,动物疫苗在纯度、效力、安全性、免疫持续时间、功效和成本方面都有所提高。第一批兽用疫苗是粗神经组织衍生产品。这些被第二代 MLV 取代。第三代涉及细胞培养中产生的灭活生物制剂,通常含有佐剂,例如氢氧化铝。第四代包括重组产品。在过去十年中,全球生产了许多不同的兽用疫苗,用于伴侣动物、牲畜或野生动物,以管理免受狂犬病病毒感染(补充表 S4)。除了宠物和牲畜外,那些在公共场所(包括宠物动物园)饲养的圈养动物也应该接种疫苗。由于没有疫苗批准用于所有处于风险中的哺乳动物,因此建议兽医批准超说明书使用。
尽管生产商和疫苗种类繁多,但所需剂量和实际生产的剂量数量不足以满足全球需求。例如,在中国,仅针对狗和猫,就需要从国内和进口部门额外接种4000多万剂。在农村地区,健康差异很明显,通过野生动物传播病毒在亚洲和非洲仍然是一个令人担忧的问题。骆驼等几种家畜的疫苗供应不足。动物保护组织和地方政府之间的伙伴关系可以提供一种解决方案,为特定的农村条件量身定制成功的兽医计划。随着未来利润丰厚的市场的出现,人们敦促警惕在兽药中非法使用假冒疫苗,如人类狂犬病生物制剂。COVID-19大流行后出现的其他担忧与发达国家和中低收入国家对疫苗的犹豫有关,而此时是需要使用现代安全、有效和免费的兽用疫苗来保护处于危险中的动物及其主人的关键时刻。除了考虑在病毒暴露之前为处于危险中的动物推广疫苗外,根据特定的产品标签要求,它们可能太年轻或未列出,并且可能由于其他原因而错过。在当地兽医法规允许的情况下,幼稚动物的PEP机会也是一种循证选择。
继续使用许可生物制剂预防生产性溶菌病毒感染部分取决于特定疫苗的质量,以及拟议疫苗的生活史阶段和暴露状态,以及兽医疫苗接种者的经验(表 3)。广泛的指导基于一般免疫学参数和最佳生物医学实践。鉴于宿主物种、传染源、环境变量和可用产品之间复杂的相互作用,始终建议与当地主题专家和政府当局合作,以遵守当地法规。
表 3.关于在家养哺乳动物中使用狂犬病疫苗来管理狂犬病病毒感染的一般指南。
| 宿主状态 | 管理考虑 |
|---|---|
| 幼稚、未暴露、健康 | 暴露前疫苗接种(初免、后期加强、根据标签建议或 1 年后) |
| 幼稚、暴露、健康 | 安乐死,或严格隔离 ~3-6 个月(如果疫苗获得此类用途许可,或获得农业和公共卫生批准,可以考虑暴露后预防管理) |
| 以前接种过疫苗,未接触过,健康 | 根据疫苗标签适应症,每年或每三年定期加强一次 |
| 以前接种过疫苗,接触过,健康 | 立即加强并观察 ~45 天,如果出现病毒性脑炎的兼容迹象,则实施安乐死 |
| 怀孕、幼稚、健康 | 暴露前疫苗接种临时 |
| 怀孕、以前接种过疫苗、健康 | ~孕晚期加强剂 |
| 新生儿,健康,从接种疫苗的母牛那里接受初乳 | ~3-6 个月时接种暴露前疫苗 |
| 新生儿,健康,来自幼稚的健康母牛 | 第一次健康检查时接种暴露前疫苗 |
| 新生儿,健康,来自狂犬病母 | 镇静水坝(然后安乐死),考虑紧急剖宫产,立即为新生儿接种疫苗(3 个月时加强针)等,并非常谨慎地使用足够的个人防护装备 |
| 任何具有兼容狂犬病临床症状的动物,无论年龄或疫苗接种状况如何 | 立即实施安乐死并适当管理可能暴露的同窝伴侣 |
9. 精选区域流行病学亮点
9.1欧洲狂犬病流行情况
尽管20世纪两次世界大战对地区产生了重大影响,但在过去200年中,非洲大陆在狂犬病病毒的诊断、控制和预防方面取得了重大进展。例如,19世纪末,狂犬病疫苗的概念起源于欧洲。经过几个世纪的流行,犬狂犬病的控制甚至在疫苗接种出现之前就开始了,主要包括流浪狗扑杀、栓养、强制戴口套和改进的卫生措施。此后,肠外犬疫苗接种迅速证明了其在控制并最终消除犬狂犬病方面的有效性。
1970年代末,欧洲是第一个对野生动物进行口服疫苗接种的大陆,为其他大陆的考虑铺平了道路。事实证明,该方法在控制整个欧盟(EU)的狐狸和貉疾病方面非常有效,2019年狂犬病已接近消除。例如,2018 年,仅三个欧盟成员国仅报告了 8 例 RABV 病例(6 例野生动物病例和 2 例家畜病例)。与 2010 年相比,这一数字大幅下降,当时 9 个欧盟成员国的病例数超过 1500 例。目标是到 2020 年实现欧盟野生食肉动物和家畜病例为零,这一目标在当时似乎是可以实现的。西欧对野生动物狂犬病的广泛控制减轻了总体农业和公共卫生威胁。今天,由于消除犬类狂犬病,欧盟的兽医疫苗接种是根据具体情况进行的。主要风险是外国旅行者和来自狗RABV持续存在的其他地区的病例转移。
与世界其他地方一样,蝙蝠狂犬病的识别始于1950年代。迄今为止,除RABV外,欧洲还发现了至少6种其他狂犬病病毒。目前还报告了5例来自EBLV的人类地方病病例,但尚未报告其他本土蝙蝠狂犬病病毒病例。1例人类DUVV病例是从肯尼亚输入到荷兰的,未接种疫苗的旅行者曾接触过非洲本土蝙蝠狂犬病。
东欧的冲突地区危及整个地区取得的大量农业、公共卫生和经济进步。乌克兰的政治事件影响疾病预防和控制(图8)。例如,摩尔多瓦、罗马尼亚和波兰的当地狂犬病情况恶化。其他消除野生中食肉动物狂犬病的周边国家(即斯洛伐克和匈牙利)最近再次感染。目前欧洲的 RABV 分布表明,任何地方的流行病学状况都非常脆弱——与邻国密切相关。在过去十年中,在其他 EID(例如,非洲猪瘟、口蹄疫、高致病性禽流感、蓝舌病、块状皮肤病等)的背景下,这种先前的成功给维持有关狂犬病病毒、狂犬病疫苗供应和局灶性疾病优先级的公众教育带来了意想不到的挑战。
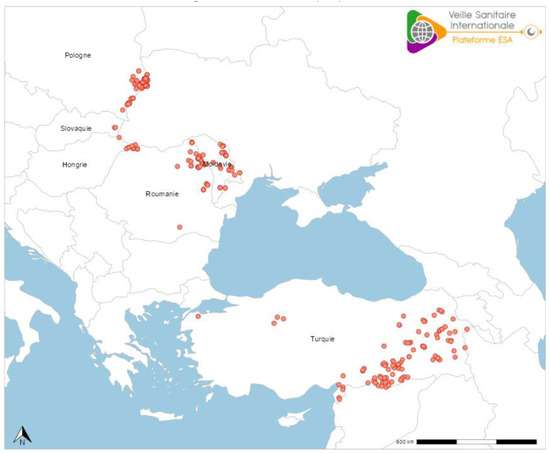
图8.显示2024年1月至2025年5月12日中欧和东欧部分地区报告的RABV病例(红圈)的地图(红圈)向动物疾病信息系统报告数据的国家(https://food.ec.europa.eu/animals/animal-diseases/animal-disease-information-system-adis_en,2025年5月19日访问)。图源:Epidémiosurveillance Santé Animale 平台(https://www.plateforme-esa.fr/fr,2025 年 5 月 19 日访问)。
备注:丽莎病毒就是广义的狂犬病病毒和狂犬病相关病毒的总称
9.2. 亚洲和印度次大陆狂犬病
动物和人类狂犬病的最大负担在于亚洲。除RABV外,该地区还记录了至少6种其他公认或推定的属于I系群的狂犬病病毒,来自中亚、中国和斯里兰卡。2007年,在俄罗斯远东地区Ozernoye村接触蝙蝠的一名20岁女性死亡病例中分离出的一种狂犬病病毒最初被称为Ozernoe病毒。后来的分析表明,这是IRKV,并且还发生了其他几例人类病例。人们对此类非 RABV 狂犬病相关病毒的更广泛分布或流行病学意义知之甚少,这凸显了当前监测和研究工作中的巨大差距。
报告和模型估计,印度、中国和菲律宾是亚洲受影响最严重的国家之一。在该地区,中国已经取得了进展,人类病例逐渐减少。1981年,有>7000例人类狂犬病感染的报道,到2014年下降至<1000。2020年仅记录了202例人类病例,分布在143个地区。继续采用“同一健康”方法表明,该国可以实现“2030零死亡”(ZBT)的目标,尽管农村地区的重点仍然是一个挑战。这种乐观情绪在其他领域可能并不现实。例如,在菲律宾,2022-2023年间报告病例有所增加,至每年350>死亡。作为一个群岛,病毒传播似乎受到海洋的阻碍,岛屿间传播很少,这可以用来改进未来的狗狂犬病控制计划。印度次大陆与中国的稳步进步或菲律宾海洋提供的潜在障碍形成了鲜明的对比。印度缺乏关于人类死亡或狂犬病动物的可靠数据,尽管实验室方法已经有了显著改进。
据信,全球狂犬病的大部分负担发生在印度。迄今为止,RABV是印度唯一有记录的病毒,家犬是主要的宿主和传播媒介。1980年,印度北部出现了一只患有狂犬病的灰头狐蝠(Pteropus poliocephalus)感染了“弹状病毒”的报道,但没有进一步的特征。最近在2013-14年期间对印度东北部那加兰邦(该地区每年可能收获数千只蝙蝠用于食用和用于当地传统医学)的监测提供了蝙蝠暴露于一些未定义的狂犬病病毒的血清学证据、。然而,一项针对阿萨姆邦蝙蝠的类似调查没有发现血清学暴露于RABV或任何其他狂犬病病毒的证据。这种血清学的动物流行病学用途可能对狂犬病病毒活性有深入的了解,但高度依赖于实验室方案和所采用的分离株。
据估计,印度每年约有20,000人死于狗传播的RABV,每年约有2000万例狗咬伤病例。虽然这些死亡中的大多数被认为是由 RABV 造成的,但其他狂犬病病毒的影响尚不清楚,因为“……这些感染很容易被遗漏,因为该病与狂犬病脑炎没有区别,狂犬病脑炎在非洲和亚洲几乎总是在临床上得到诊断。
印度国内的负担并不奇怪,因为该国自由漫游的狗数量异常多,估计超过 6000 万只。在这种环境中,这些“自由漫游”的狗不一定是传统意义上的“流浪狗”,因为大多数狗都松散地隶属于社区,使它们成为“社区拥有的”狗,而不是真正的“无主”狗。此外,即使是被认为是“拥有”的狗通常在大部分时间里自由漫游,因为在这些环境中养狗的概念不一定扩展到常规疫苗接种、预防性医疗保健、节育或坐月子(图 9)。整个印度次大陆大规模自由漫游和未接种疫苗的狗种群中狂犬病的元种群动态,加上自由漫游的狗与人类、家畜和野生动物持续的、无人监督的互动,对狂犬病控制计划提出了重大挑战,这些计划依赖于双重大规模疫苗接种和绝育作为“动物节育”(ABC-ARV)的主要手段。

图9.与许多中低收入国家一样,印度的自由放养狗数量众多,并且依赖当地社区的食物和住所来源。
从历史上看,大规模狗疫苗接种(mass dog vaccincin, MDV)计划一直是近期控制和消除犬狂犬病的成功努力的支柱,从而在区域层面减少人类狂犬病死亡。例如,三十多年来,拉丁美洲和加勒比地区国家每年总共为4000多万只狗接种狂犬病疫苗。这是由于泛美卫生组织(PAHO)协调的大陆规模犬狂犬病疫苗接种运动的坚定政治承诺。然而,在没有强有力的政治承诺的情况下在印度实施类似的 MDV 战略带来了重大挑战。这些犬类种群的周转率很高,这是导致每年疫苗接种活动之间疫苗接种覆盖率持续降低的原因。此外,为了有效阻止RABV的传播,理想情况下,至少70%的区域犬种群应在短时间内(例如,一个月)接种疫苗。此外,控制RABV疫情不仅需要MDV,还需要对疑似狂犬病动物进行安乐死,以及局灶和围区疫苗接种以遏制病毒传播。然而,在印度,2001年制定的绝育和抗狂犬病疫苗接种(ABC-ARV)政策在一些领域被解释为禁止杀死疑似狂犬病的狗。据估计,印度有 8500 万只狗,其中超过 70% 是自由漫游的,在 ZBT 的时间戳内实现持续和协调的疫苗接种覆盖率是一个值得称赞但无法实现的目标。
目前的ABC-ARV计划旨在通过减少易感犬的数量和限制公狗行为的某些方面(如局部传播和打架)来减少犬种群更替并减少RABV传播。在现实世界中,这些ABC-ARV项目往往无法实现犬种群规模的预期减少和必要的狂犬病疫苗接种覆盖率。例如,使用 DogPopDY(一种基于代理的建模工具,即使在理想条件下(例如所有狗都平等且易于接近的封闭种群)模拟真实的计算机狗种群,ABC-ARV 计划也未能达到阻断 RABV 在狗种群中传播所需的疫苗接种覆盖率(图 10)。这种失败主要是由于印度自由漫游的狗种群规模庞大,阻碍区域覆盖的后勤挑战,以及最关键的是,长期以来缺乏总体强有力的政治承诺。
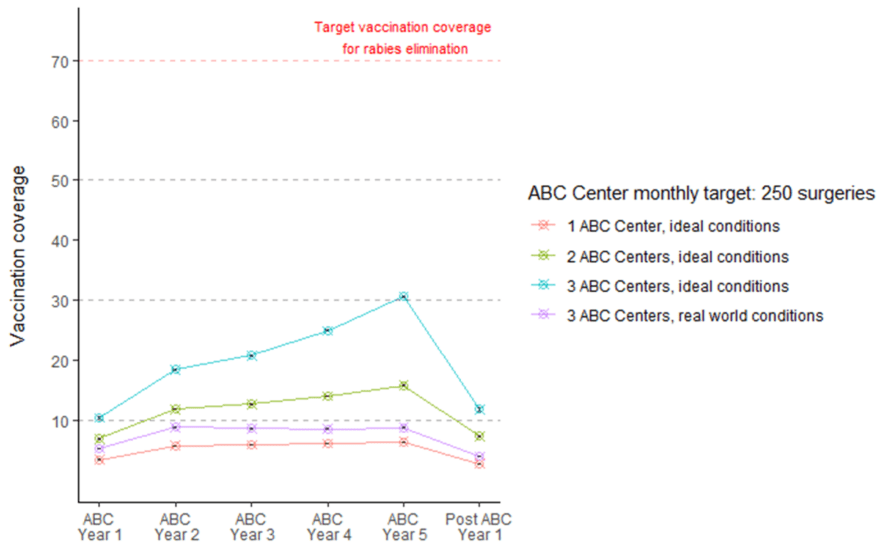
图 10.在印度,即使在理想条件下,目前的 ABC 计划也无法实现/维持狂犬病控制所需的疫苗接种覆盖率。
作为印度 RABV 的主要宿主和载体,家犬也是病毒传播给其他家畜的主要来源。在整个印度,自由放养的牛在城市和农村都无处不在。狗介导的溢出效应对印度牛(超过25个独特品种)并不少见,尽管它可能被低估了[333,362]。在农村地区,疑似患有狂犬病的牛(通常是在已知的狗咬伤事件之后)有时会在市场上卖给毫无戒心的买家。这进一步引发了人们对通过非咬伤暴露将病毒传播给人类的感知风险的担忧,包括食源性传播的可能性,尽管这种情况很少见。正如弗莱明 (1872) 在历史上所指出的那样,……“很大一部分证据支持来自奶牛或其他生物的牛奶在受狂犬病影响时是无害的……”,但仍然安全地认为“……必须永远记住,一个积极的事实抵得上一千个消极的事实……”。虽然没有书面证据证实RABV通过食用受感染动物的肉类或奶制品传播,但理论上受感染牛的生奶(未经巴氏消毒/未煮沸)的传播风险经常引起媒体的关注和公众的关注。一些个案报道存在,包括最近在印度报道的一例病例,涉及一名女性在食用未经巴氏消毒的牛奶后出现狂犬病。
由于狗和其他动物的多种接触源,人类病例很常见,尽管印度在 2005 年停止了用于现代生物制剂的神经组织产品。遗憾的是,用于PEP的现代生物制剂(即狂犬病疫苗、RIG、狂犬病单克隆抗体)并不容易获得,而且往往供不应求。此外,这些环境中的许多医疗专业人员对适当的PEP了解不太理想。狗咬伤受害者或其看护人通常来自偏远农村地区,通常必须进行名副其实的“探险”以寻找 PEP,从而导致 PEP 延误甚至不给药。为了帮助解决PEP依从性差的问题,交互式Web应用程序ZeroRabiesApp(ZRA)提供了一种用于治疗狗咬伤病例的即时工具。ZRA 可通过智能手机和电脑免费访问(https://anyadoc.shinyapps.io/ZeroRabiesINDIA/,2025 年 5 月 1 日访问)。该工具提供对世卫组织和美国免疫实践咨询委员会制定的最新指南的访问。它根据用户提供的咬伤日期(暴露日)生成定制的 PEP 时间表,该时间表可以与用户的医疗保健提供者共享。ZRA 还提供对狂犬病生物制剂数据库的访问,该数据库可用于查找 PEP 产品目前库存的最近地点,确保更及时地获得必要的护理。
9.3. 非洲狂犬病流行情况
尽管自20世纪中叶以来就有了有效的预防措施,但估计每年仍有数万人死亡,其中95%的狂犬病病例在非洲,仅次于亚洲。虽然全球主要关注通过大规模疫苗接种、人类PEP和公众意识运动来消除犬介导的狂犬病,但其他溶疮病毒在很大程度上被忽视了。不同宿主和病原体的多重、交织的复杂性普遍存在,影响了疾病监测和控制,包括雄心勃勃的 ZBT 目标。虽然重点关注RABV,但在整个非洲大陆,存在一种更具包容性的方法,即所谓的“狂犬病相关病毒”(表1)在1950年代首次被发现。
有效的动物狂犬病控制依赖于强大的监测系统和有针对性的干预措施。在整个非洲,很少有历史例子可以效仿,而且往往依赖于公私合作(例如坦桑尼亚)。在南非夸祖鲁-纳塔尔省等地区,包括培训、意识、MDV、疫苗库和可及的 PEP 在内的综合策略导致人类狂犬病发病率暂时下降。然而,一旦国际支持消失,进展就很短暂。在许多非洲国家,监测系统仍未开发,导致报告不足和控制不足。加强这些系统对于准确收集数据和制定有效的应对策略至关重要。
ZBT 倡议于 2015 年启动,旨在到 2030 年消除犬介导的人类狂犬病死亡 。ZBT 战略强调 MDV、公众意识和可及的人类 PEP。基本概念是合理的,但时间表过于雄心勃勃。面对非洲大陆的有限进展,挑战仍然存在,尤其是在资源和基础设施有限的地区。实现 ZBT 目标需要优先考虑、持续承诺、可靠的资金和国际合作。对消除犬类狂犬病的高度关注导致了研究和资源分配的差异。所有其他 lyssaviruses 的研究较少,导致理解和准备有限。这种不平衡在整个非洲都很明显,犬狂犬病在那里流行,而其他狂犬病病毒则很少被研究。解决这一差距需要一种更具包容性的方法,在优先控制犬类狂犬病与忽视“同一个健康”领域的其他狂犬病病毒和对 EID 的担忧之间取得平衡。非洲承受着沉重的狂犬病病例负担,据估计,非洲大陆约占全球相关死亡人数的35%-42%。除 RABV 外,此类疑似人类死亡中有多少比例是由 RABV 以外的其他 lyssavirus 引起的尚不清楚,因为大多数病例未通过实验室检测确诊(或表征)。相对于家养动物和野生动物案件,情况也是如此。
9.3.1. 碎片化的控制程序
多个非洲国家开展了许多分散的狂犬病控制计划。与欧洲或美洲的情况不同,大多数国家没有通过国家狂犬病集中控制计划(NRCP)进行协调。这种缺乏集成会导致效率严重低下。没有统一的系统来监测和评估预防和控制工作。因此,疫苗接种和教育活动可能会在已经覆盖的社区重复,从而导致资源和努力的浪费。缺乏指定的 NRCP 使情况进一步复杂化,阻碍了对进展的有效跟踪,并阻碍了重复的卫生举措的成功。建立 NRCP 在一定程度上通过将包括非政府组织在内的各种利益相关者的活动整合到一个统一的战略下来应对这些挑战。这种整合有助于全面规划、资源分配以及监测和评估框架的开发,以评估控制措施的影响。
除了政府成员之外,非政府组织通过提供资源、专业知识和社区参与,在控制方面发挥着至关重要的作用。然而,如果没有协调,他们的工作就会重叠,导致程序效率低下。为了使狂犬病控制计划有效,各国必须承担起责任并领导这些举措。至少,这种领导力涉及制定国家战略、获得资金和促进所有利益相关者之间的合作。此外,还存在对国际援助的高度依赖,从长远来看,这是不可持续的。例如,美国政府最近减少的国际援助凸显了仅依赖外部支持基本卫生计划的脆弱性。虽然非洲疾病预防控制中心在广泛加强疾病监测和提供技术援助方面发挥着关键作用,但在国家和地区层面都需要加大投资和制定更具凝聚力的战略。区域合作至关重要,因为邻国必须合作防止狂犬病和其他疾病的跨境传播。一些亚洲国家已经成功地实施了综合监测和报告系统,以加强狂犬病控制工作。例如,中国湖南省和越南建立了综合系统,整合了人类和动物卫生部门的数据收集,促进了更好的监测和响应。此外,印度已开始使用标准化报告格式,以帮助确保更准确的监测和及时干预(https://rabiesfreeindia.mohfw.gov.in/,2025 年 5 月 1 日访问)。然而,非洲在很大程度上缺乏这种综合系统,那里的监测仍然分散且不一致,阻碍了有效跟踪和控制狂犬病的能力。
9.3.2. 实验室监测的挑战
薄弱的监测系统严重阻碍了非洲狂犬病的检测和了解。例如,一项涉及非洲 54 个国家中 49 个国家的研究发现,在 23 名受访者中,至少有 16 名 (70%) 的监测系统无效。虽然大多数病例被认为是由 RABV 引起的,但诊断能力不足导致所有 lyssavirus 感染的总体漏报。这种缺陷导致对人类负担的估计不精确,并掩盖了该疾病的真实流行病学。财政限制进一步加剧了监控限制。在马达加斯加,资金不足阻碍了对可疑动物的获取、适当的约束、镇静和安乐死、脑样本的收集以及必要的生物安全措施的实施,导致严重的漏报[379]。这些缺陷阻碍了有效监测,凸显了加强监测系统、增强诊断能力和为评估狂犬病实际状况提供充分支持的必要性。即使面临这些限制,实际的疾病控制和内省的病原体发现也不是相互排斥的,正如同时进行的狂犬病预防和流行病学监测所见,重点是南非境内的几种狂犬病病毒(例如DUVV、LBV、MOK、RABV等),包括在多种野生中食肉动物中鉴定RABV宿主,包括土狼、狐狸、豺狼、猫鼬等。
9.3.3. 食用受感染动物?
患有狂犬病的动物不应故意进入食物链。虽然牲畜通常是终末宿主,但患病的动物被屠宰并进入当地市场。此类事件会通过唾液污染开放性伤口和在准备供人类食用的动物时接触受感染组织而产生病毒传播的风险。这些做法使监测和响应变得复杂,因为屠夫经常因其经济价值而避免对动物进行测试,从而进一步阻碍了准确的疾病预测。虽然接触受感染的动物产品不是生产性感染的常见途径,并且适当的烹饪/巴氏杀菌可以灭活组织中存在的溶菌病毒,但在极少数情况下,如果动物处于狂犬病晚期,摄入可能会造成风险,从而产生公众焦虑,需要进行健康评估和获取有限的 PEP 。此外,除了肉类之外,还有许多关于个人食用生牛奶的报道。在津巴布韦,牛主可能不知道人畜共患病。如果被狂犬病狗咬伤的奶牛喝了牛奶,而奶牛死于狂犬病,村民可能会因为担心潜在的病毒暴露而受到监视。仅凭信息可能不会超过冒险的传统做法。埃塞俄比亚的一项调查表明,>75%的受访者对人畜共患疾病传播有良好的看法,但食用生牛奶或生/未煮熟肉类的做法仍然很高。同样,几内亚的调查参与者可能已经理解他们每天所承受的危险,基于对依赖动物的生计的价值观,但对不确定的世界表达了一种无能为力的感觉,强调“……人畜共患疾病预防需要对影响预防行为的因素有透彻和文化上的细致入微的了解“。除了家畜之外,食用丛林肉在非洲也是一种普遍的做法。在马拉瓦的一项调查中,至少有50%的受试者食用蝙蝠肉,这表明某些农村社区经常存在潜在的人畜共患风险。
9.3.4. 有病毒暴露风险的兽医
接种疫苗并担任患病动物护理人员的当地兽医有接触病毒的风险。不幸的是,关于接受 PrEP 的非洲兽医百分比的数据有限。尼日利亚伊巴丹大学的一项研究发现,15.4% 的兽医学生接受了推荐的 PrEP 方案。大约 19% 的人只接受了单剂。这表明,虽然一些专业人士对狂犬病的认识可能相对较高,但PrEP的实际接受率仍然很低,这可能是由于获取、成本和认识等障碍。如果疫苗接种不能在中低收入国家内更广泛地普及和免费提供,那么期望兽医专业人员继续从事相关的疾病监测和控制实践可能是不现实的。新的简化剂量节约方法,规定在一周内接种疫苗,而不是3周或更长时间,可能为全球关键专业人员提供更多机会。
9.4. 北美、中美洲和南美洲狂犬病
美洲是唯一一个仅存在狂犬病病毒(RABV)这一种lyssavirus病毒的地区,其本土病毒变异株的数量多于已确认的lyssavirus病毒种类。犬类狂犬病防控计划始于20世纪20年代,北美地区逐步消除了犬类狂犬病的传播。中美洲和南美洲也在取得越来越多类似的进展。目前,犬类狂犬病病毒的流行灶分布在玻利维亚、海地、委内瑞拉,最近在秘鲁也出现了相关病例。除犬类外,狂犬病病毒的主要储存宿主还存在于野生动物中,包括野生中食肉动物、非人灵长类动物(如巴西的狨猴),以及多种食果、吸血和食虫蝙蝠。值得注意的是,存在从蝙蝠狂犬病病毒向中食肉动物的病毒溢出感染和宿主转换现象。加拿大和美国均采用口服疫苗来控制郊狼、狐狸或浣熊中的狂犬病。 在加勒比海岛屿上,狂犬病出现在古巴和格林纳达(主要发生在獴和蝙蝠中)、伊斯帕尼奥拉岛(如通过犬类和獴传播)、波多黎各(如通过獴传播)以及特立尼达岛(通过吸血蝙蝠——圆耳蝠传播)。除加强监测外,抗原分型和分子特征分析有助于了解不同宿主中狂犬病病毒变异株的多样性,例如在美国这样的国家,即使不考虑蝙蝠中更多的变异株,多种中食肉动物中也存在丰富的变异株。
9.5. 加勒比地区,特别提及特立尼达:牛、小型反刍动物与水牛
在全球范围内,狂犬病导致的牲畜死亡估计为每年>5亿美元,在依赖牲畜经济的国家损失较大。虽然对这些国家的影响是巨大的,但这种损失仅占狂犬病全球经济负担的6%。动物宿主的地理差异需要采取更量身定制的策略来防止溢出到牲畜种群中。牲畜被认为是终末宿主,但却充当病毒活动的哨兵。在新大陆,特立尼达(加勒比海唯一现存吸血蝙蝠的岛屿)等国家自1930年代以来一直在处理狂犬病和牲畜问题,当时吸血蝙蝠被认为是宿主。当时,鉴于狗仍然是拉丁美洲的主要宿主,管理牲畜种群大规模爆发还没有先例。犬狂犬病主要作为公共卫生问题(例如在伊斯帕尼奥拉岛的加勒比海地区),病毒主要溢出到伴侣动物和人类身上。或者,来自吸血蝙蝠的 RABV 主要传播给牲畜。因此,这种新大陆现象被视为动物生产问题,对当地粮食安全构成潜在威胁,特别是对小农户而言。幸运的是,由于最近拉丁美洲疾病范式的转变,人们更强烈地关注食草动物的狂犬病。食草动物狂犬病的风险人群比犬传播的RABV风险人群更窄。考虑到兽医和其他动物卫生工作者应该进行 PrEP,这些人主要包括农民和屠夫。因此,与中食肉动物和蝙蝠相比,牲畜对人类溢出感染的风险较低。事实上,在过去5年中,只有1例人类病例报告归因于美洲家养食草动物的病毒传播。该病例发生于2023年,来自巴西米纳斯吉拉斯州的一头牛。然而,随着宿主种群的扩张(例如气候变化),更多的食草动物与宿主相互作用(例如,吸血蝙蝠捕食增加)可能会增加这种风险,更多的狂犬病食草动物会产生机会性溢出效应。以吸血蝙蝠为媒介,牲畜可以充当人类攻击的缓冲器。然而,如果牲畜数量减少,人类可能会成为与公共卫生直接相关的替代宿主。
在特立尼达,由于多种因素(例如,农业激励措施、市场需求、土地利用变化等),畜牧业的概况随着时间的推移而发生了变化。随着这一变化,动物的狂犬病风险状况也发生了变化。例如,从1971-2015年,小型反刍动物病例明显增加,尤其是山羊。因此,狂犬病控制措施发生了变化。山羊的疫苗接种是在特立尼达与绵羊一起进行的,因为由于吸血蝙蝠袭击的增加,它们现在被认为是感染狂犬病的高风险物种。这些较小的食草动物可能会证明报告偏差,因为它们要么更容易检索以进行测试(阳性偏差),要么更容易丢弃(负偏差)。
水牛(Bubalus bubalis),当地称为“野牛”(不同于北美本土野牛)或“猪牛”,在1905-1908年和1923-1949年期间的两次重大事件中从印度进口到特立尼达。他们被带去协助糖和椰子行业的工作,拖着手推车和犁地。进口品种包括沼泽型和河流型动物,其遗传种群被选择性地用于生产“水牛”,这是一种独特的特立尼达杂交种,专门用于肉类生产和吃水。这些动物在全岛都很常见,直到 2003 年甘蔗产业低迷。此后,水牛在二十年内下降了58%。到2012年,这一数量进一步减少了25%,全国有1153只动物,其中大多数在三个国有农场,其中最大的农场受到布鲁氏菌病高患病率的威胁。RB51布鲁氏菌病疫苗对牛有效抵抗流产双歧杆菌感染和流产,但尽管进行了血清转化,但未能在特立尼达水牛中提供类似的疗效。RB51疫苗接种后缺乏细胞介导的免疫应答可能是缺乏疗效的原因。狂犬病疫苗对多种牲畜非常有效,尽管它们尚未获得许可或验证可用于杂交物种。然而,鉴于 RB51 疫苗在水牛中的功效存在显着挑战,有必要对该物种的狂犬病疫苗接种提供的免疫覆盖率进行比较研究。
在大型食草动物中,水牛的狂犬病报告频率低于牛。美洲的特立尼达、墨西哥、巴西、厄瓜多尔和秘鲁以及埃及、印度和中国的报告较少。水牛狂犬病病例的总体检出有限可能是由于多种因素共同作用造成的。对于由食肉动物(如狗)传播的RABV,这些强壮的动物被认为更有能力抵御地面攻击。尽管它们在物理上抵御蝙蝠的空中攻击的能力较差,但它们具有一些解剖学优势,可以减少这些体型小得多的哺乳动物的咬伤。例如,水牛和牛的皮厚度存在显着差异。水牛在吸血蝙蝠通常以觅食为目标的区域(即颈部、肋骨、背部、飞节和尾基部)表现出明显更厚的皮肤。这些区域会随着年龄的增长而增厚。这些特征可能会妨碍吸血蝙蝠轻松进食(即咬人)。然而,吸血蝙蝠在地面上打滚在泥土或水中时,通过咬住水牛较软的鼻孔皮肤(图11)来以水牛为食,这是巧妙的聪明才智。

图 11.吸血蝙蝠可以咬伤水牛的鼻孔并在鼻孔上造成病变。
其他可能导致水牛病例报告减少的因素包括监测偏差和动物相对难以进行观察和测试,尤其是牧场上的动物。在特立尼达,通常在如此广泛的管理条件下饲养的水牛只有少数大型农场。个体农民的财产通常每个农场只有2-5只动物。许多农民实行畜牧或“无地”农业,食草动物从一个地方转移到另一个地方放牧。在特立尼达,动物在进食期间有时会在道路上自由漫步,这给车辆带来了安全问题(图 12)。岛上的另一个情况是从南美大陆非法进口牲畜。报告谴责大量“流浪”动物在该国南部的村庄和城镇周围徘徊。由于担心诉讼或迫害,此类动物的主人很少站出来报告生病或垂死的动物或要求接种狂犬病疫苗。

图 12.特立尼达道路沿线的自由放养牲畜。
虽然从理论上讲,粗放或畜牧动物饲养增加了野生动物宿主与牲畜的相互作用并增加了病毒传播的机会,但随之而来的是,它限制了直接观察动物的机会,从而减少了病例检测的机会。因此,由于监测和报告的局限性,在这种做法盛行的地区,牲畜狂犬病的真实患病率可能被严重低估。广泛饲养牛群管理面临的其他挑战包括动物识别、狂犬病疫苗接种的捕获和限制以及临床参与方面的困难。有限的检测和报告也可能出现病例误诊可能仅基于临床观察。兽医的参与可能不是特别频繁。与牛相比,水牛的潜伏期可能变化更大(2-12周)。临床症状基本相似(例如,异食癖、磨牙症、凝视、大量流涎、嘶哑吼叫、后肢麻痹、不协调、瘫痪、卧位和发病后 5 天内死亡)。然而,在其他牛科动物中,水牛中明显出现尾巴麻痹或尾巴扭结的记录尚未常规。没有狂犬病疫苗获得水牛的许可,尽管这些生物制剂被认为在各个物种中都非常有效。无论如何,由于这些动物独特的体格,管理带来了与这些高度攻击性动物的约束和疫苗接种技术(例如针头位置和长度)相关的后勤挑战,以成功分娩。
9.6. 澳大利亚地区狂犬病
从历史上看,大洋洲(主要由水组成,有数千个火山岛或环礁)通常被认为是“无狂犬病”的(但没有实施常规的实验室监测)。然而,即使在那里,流行病学情况也并非一成不变,正如关岛观察到的那样,1967-1968年期间疾病传入和控制。此外,除了狗之外,患有狂犬病的野生动物,如蝙蝠,也可能被转移到遥远的岛屿。
太平洋地区的这种罕见性、源头和汇之间的相对“出色隔离”、从1866-1867年期间第一次也是唯一一次可疑的“爆发”开始的漫长时间范围,以及严格的隔离措施的实施,似乎使澳大利亚相信狂犬病不存在。在将那片大陆与当时美国的动物园情况进行比较时,Skerman (1964) 沉思道,显然“……建立永久感染宿主的条件不存在……”。关于从东南亚地区进口狂犬病的合法风险,一些人认为“……如果说它不能在这里发生……“,那就太愚蠢了,与”……我们最多只能说,它不必在这里发生。引入是有道理的,因为如果狗进入并成为地方性,研究人员估计>35,000次咬伤可能需要PEP。从旅行医学的角度来看,专业人士警告说,“……人类狂犬病迟早会发生……通过在另一个国家感染病毒的人抵达澳大利亚。关于这种“遥远的可能性”(如器官移植病例)的问题,一些医生开始关注这个话题,因为“……类似狂犬病的脑病很少见,应该考虑所有可能导致不明神经系统疾病的原因。
1987年,一名10岁的澳大利亚男孩在死后4个月被诊断出患有狂犬病,这有点先见之明。他曾与母亲一起游历亚洲(发病前 16 个月,可能在印度被猴子咬伤后被感染),回国后约 8 个月生病。显然,这个独特的案例表明“……需要将狂犬病纳入脑炎的鉴别诊断……“,同时理解”……确定狂犬病的诊断可能很困难。此后,第二例病例在死后 3 个月被诊断出来,发生在一名 10 岁的越南女孩身上——距她被推测在国外暴露6年多。正如 Grattan-Smith 所说,“……狂犬病发生在澳大利亚,在急性脑炎的鉴别诊断中需要考虑……”。慢慢地,即使在这些不寻常的人类病例之后,也只有少数人相信“它可能在这里发生”,尽管显然仍然关注犬狂犬病对外构成的明显主要威胁,但实际上不是来自内部的。
地方性狂犬病的发现仅通过不相关的 EID 事件发生。1995年,马病病毒(又名今天的亨德拉病毒)在马中爆发(并伴有相关人类病例)后,澳大利亚调查人员开始寻找与疑似野生动物中病毒出现有关的明显宿主。作为流行病学调查的一部分,1996 年,新南威尔士州巴利纳的一只幼年黑狐蝠 (Pteropus alecto) 在被发现在一棵无花果树下无法飞行后被安乐死。该物种是澳大利亚最大的蝙蝠,栖息在由数百到数千只个体组成的明显“营地”中(图 13)。尸检显示为严重的非化脓性脑炎。从组织学上讲,神经元内存在嗜酸性粒细胞、胞浆内包涵体。中枢神经系统的电子显微镜图像显示细胞体细胞质内的病毒核衣壳聚集体。在大脑内,狂犬病的免疫组织化学测试呈阳性,1995 年回顾性蝙蝠中枢神经系统也有类似的发现,具有多个代表性的胞浆内包涵体(补充图 S4)。

图 13.黑色狐蝠 Pteropus alecto 的艺术描绘。(由 J. Gould 提供,澳大利亚哺乳动物,1863 年,维基共享资源,https://commons.wikimedia.org/wiki/File:Pterogus_alecto1.jpg,2025 年 5 月 19 日访问)。
在实验室小鼠体内通过受感染组织后,通过电子显微镜检查脑匀浆,发现弹状颗粒,这是弹状病毒的典型特征。系统发育分析表明,与蝙蝠相关的病毒与经典RABV密切相关。使用一组 MAb 进行抗原分型证实蝙蝠病毒分离株是一种裂病毒,与该属的其他病毒不同,但与 RABV 非常相似。
蝙蝠病毒分离株被抗血清中和RABV。该病毒暂时被称为“翼状裂虫病毒”,在被称为 ABLV 之前。除了在P. alecto中诊断外,在昆士兰州北部的小红狐蝠(Pteropus scapulatus)中也发现了ABLV。蝙蝠的发病率和死亡率涉及多种因素,如毒性,因此,对于断诊,需要进行特异性诊断,而不仅仅是依赖临床症状或组织学检查。进一步的系统发育分析支持了ABLV是Lyssavirus属的新成员的观点。
随着对蝙蝠的监测增加,以更好地了解ABLV的动物流行病学,先前针对旅行者的预防建议不断发展,以帮助保护国内暴露个体的健康。渐渐地,其他关于蝙蝠 ABLV 的报道增加到包括北领地、昆士兰州、新南威尔士州和维多利亚州。最初的大多数蝙蝠采样都是机会主义的,并且集中在澳大利亚东部。为了检测先前死于狂犬病的病例,一项回顾性流行病学研究回顾了 1992 年至 1996 年北领地不明原因的人类脑炎病例。目的是检测任何可用的临床标本以寻找狂犬病病毒感染的证据,并调查临床医生对诊断性检测的使用情况。不幸的是,只剩下两个样本(均检测呈阴性)。该审查发现,在脑炎病例中存在相当一部分不明原因的疾病,但无法排除受影响人群中涉及 ABLV。由此产生的建议建议,临床医生应检测脑炎患者的狂犬病病毒感染,在人类死亡原因不明时进行尸检,并适当储存任何标本以进行 EID 检测。
在1996年发现后数月内,一名39岁的昆士兰州妇女报告了首例由ABLV引起的人类狂犬病病例,她在从事蝙蝠护理工作期间感染蝙蝠后死于脑炎。黄腹袋蝙蝠或鞘尾蝙蝠 (Saccolaimus flaviventris) 咬了她,而不是来自翼形蝙蝠,显然是一种食虫性栓塞动物物种(Saccolaimus flaviventris)。接触大约一个月后,她的表现类似于经典狂犬病(有点奇怪的是,神经学报告将该病例命名为“来自果蝠的非狂犬病裂解病毒人类脑炎:澳大利亚蝙蝠裂伤病毒(翼状裂伤病毒)感染”)。患者在发病后~20天死亡。尽管最初认为这种人类分离株的序列与从翼形蝙蝠中获得的序列最相似,但后来被确定为与狂犬病病毒相关的变异。鉴于日益严重的公共卫生问题,1997年,为澳大利亚接触蝙蝠的人提供狂犬病PEP以及为持续接触蝙蝠的人提供PrEP与在国外接触RABV的旅行者保持一致。然而,在初步承认 ABLV 后,西澳大利亚州公众对预防的需求仅略有上升。
1998年,昆士兰州一名37岁女性发生了第二例死亡。1996 年 8 月,即确诊前 27 个月,这名妇女正在参加户外烧烤。一只狐蝠落在一名儿童的背上,这名妇女在移开蝙蝠时被咬伤。在 48 小时内,她从医生那里接受了抗生素和破伤风类毒素(但不是 PEP!?六个月后,她询问了“蝙蝠病毒”的检测。尽管被建议接受 PEP,但她拒绝了。她于 12 月去世,即发病 19 天后。她的“……临床表现、持续时间和疗程……与狂犬病中观察到的几乎没有区别,尸检组织学发现也是如此,包括Negri尸体。从她的唾液中产生的扩增子的初始序列表明与 ABLV 相关的感染,但与当时参考实验室获得的其他序列有些不同。随后对细胞培养传代后获得的CNS分离株的分析证实了由ABLV的翼龙变异引起的感染。经过漫长的潜伏期和最终诊断,孩子和聚会上的其他暴露者接受了 PEP。随着耸人听闻的媒体头条大肆宣传“杀手蝙蝠”,公众对预防的需求急剧上升。
一项回顾性研究发现,根据对其他狂犬病病毒感染的检查,ABLV引起的中枢神经系统原发性病变与经典RABV相同。持续的监测支持了这样一种观点,即澳大利亚各地的食果蝙蝠和食虫蝙蝠都是潜在的ABLV宿主,尽管实验室对提交的蝙蝠感染的检出率仍然很低,正如全球蝙蝠狂犬病监测所预期的那样。关于延续,在翼龙群体内,特别是“……狐蝠通常聚集(和互动)的大型季节性游牧、多物种群落为 ABLV 的物种间和区域间传播提供了机会……”。
由于 ABLV 而对此类狂犬病病例的承认继续给公共卫生和野生动物保护带来挑战。例如,将一只自然感染的幼年狐蝠放入圈养康复中,随后在持续护理期间接触未接种疫苗的人,消除了提高社区和专业对该疾病及其相关风险的认识的需要。正如预期的那样,兽医被纳入接受PrEP作为预防措施的更广泛的高危人群。越来越明显的是,ABLV感染引起的基础疾病及其主要诊断、预防和控制方法与RABV和其他I型溶血病毒在蝙蝠体内没有本质区别。此外,有人怀疑,尽管ABLV在20世纪末才被发现,但在欧洲殖民之前,ABLV很可能在澳大利亚存在了几个世纪的动物性动物。进一步的研究表明,ABLV 与 RABV 一样,受到“……强选择性约束,…宿主物种、细胞趋向性和生态条件的稳定性。新的管理策略建议对蝙蝠进行疫苗接种,但此类建议从未实施。尽管有令人信服的客观数据相反,但一份有些令人困惑的公共卫生情况说明书指出,“……狂犬病是一种能够咬人和抓挠的哺乳动物的感染。虽然它发生在世界许多地方,但在澳大利亚却没有发生……”。
除了两人的死亡外,1995 年至 2005 年的所有 ABLV 感染病例仅在蝙蝠中检测到。溢出到其他动物的可能性被认为很低。在全球范围内,大多数与蝙蝠狂犬病病毒相关的狂犬病病例都是种内病例(图 4)。尽管RABV的宿主范围很广,但与狂犬病中食肉动物感染相比,从蝙蝠到狗和猫的溢出感染病例并不常见。在澳大利亚,在实验室条件下对少量猫和狗进行药敏试验,结果仅对单一ABLV分离株产生有限的非致命性感染。澳大利亚狗和猫接触蝙蝠后的公共卫生建议包括密切观察3个月。2000 年至 2005 年的监测发现,只有 5 例狗与狂犬病蝙蝠有过接触。所有这些狗在 90 天后都保持正常。然而,2004 年,一只狗在 1 月份接触了一只未经测试的蝙蝠,在 4 月份表现出剧烈的行为变化,并野蛮地袭击了一名儿童。这只狗在没有经过测试的情况下就被摧毁了。非常谨慎地,孩子接受了 PEP。在发现 ABLV 后的十年内,对人类预防的积极审查导致修改了开始疫苗接种的时间。由于诊断性检测的可靠性,在等待结果期间延迟长达48小时被认为是可以接受的。一项对2005-2007年悉尼地区PEP实践的回顾发现,患者依从性合理,但给药并不总是在规定的时间内进行。
与其他嗜神经性病因(例如疱疹病毒)相比,人类狂犬病很少见,但应纳入人类脑炎的鉴别诊断。然而,此类案件的解决仍然是一个挑战。一项关于1979-2006年澳大利亚脑炎相关死亡的全国性综述发现,不明原因死亡的比例从1979-1992年的47%上升到1993-2006年的57.2%。据推测,从 1998 年到 2010 年,ABLV 缺乏额外的人类病例被认为与对暴露个体实施更广泛的 PEP 措施有关,尽管这种狂犬病病毒被认为“……随着澳大利亚人口密度的增加,以及因此人类与狐蝠之间的互动水平的增加,它始终在澳大利亚人的健康中发挥着重要作用。。向专业蝙蝠护理人员和志愿者提供公共教育和提供 PrEP 是健康促进的重要组成部分。对2007-2011年新南威尔士州接受PEP的个体进行的横断面分析显示,在澳大利亚,大多数接触者是试图营救蝙蝠的人。
2013年,昆士兰州一名8岁男孩发生了第三例人类病例。2012年11月,在圣灵群岛度假时,他的前臂被狐蝠抓伤。尽管公共卫生部门广泛宣传了蝙蝠接触的危险以及在接触蝙蝠时需要立即进行医疗评估,但他没有收到 PEP。显然,他的兄弟姐妹和朋友知道这件事,但他的父母却不知道。接触后大约 8 周,他的发烧、厌食、疼痛和行为改变越来越严重,住院治疗。根据血清学和半巢式 RT-PCR,狂犬病狂犬病病毒的阳性结果发生在入院第 14 天。基因测序显示与翼状动物 ABLV 匹配。他的临床课程“……遵循脑炎狂犬病的模式……”他于2013年2月去世,即发病28天后。令人担忧的是,2011-2013 年在新南威尔士州进行的一项调查发现,参与者“……报告说,人们普遍缺乏对 ABLV 的认识,尤其是疾病风险……“。
同样在2013年,家畜报告了两例病例,两匹未接种疫苗的马都在同一围场感染,可能是由于接触了一只受感染的蝙蝠。在那次通讯中,提到了具有类似临床表现的轶事病例的报告,但既没有考虑 ABLV,也没有进行实验室测试。自这两起不相关的死亡事件以来,迄今为止,非蝙蝠中没有报告更多的 ABLV 病例。提高公众意识、向农村社区进行外展、改善获得 PrEP 和 PEP 的机会、遵守野生动物保护的最佳做法、传染病专家和兽医的继续专业教育以及应用监测和研究是在“同一健康”背景下预防 ABLV 感染的一些关键的相互关联措施 。
回想起来,25 年前澳大利亚的一些狂犬病最终受到重视,不同的蝙蝠宿主维持了 ABLV 的不同病毒变体,这引发了有趣但悬而未决的问题。在过去的几十年里,是否有任何人类病例未被识别?土著原住民与野生动物(或蝙蝠(或班古对尤因人))之间有什么关系?在教育推广和检测方面,健康差异是否明显?本地有袋动物易感吗?对其他类群的监测工作如何,包括高度易感但引进的哺乳动物,如红狐?最初限制家畜疫苗接种是否有理由?是否有一个不明显的病毒物种屏障来限制猫和狗的溢出病例,无论多么罕见?疫苗接种状况是否最大限度地减少了免疫学初治的兽医及其工作人员参与疑似脑炎和尸检表现的患者评估?如果这种疾病被更公开地讨论为“狂犬病”,会有更多的兽医接受 PrEP 吗?考虑到客观标准,经常被引用的所谓“狂犬病样”疾病是什么?对亨德拉病毒的强调是否给普通民众带来了对狂犬病、蝙蝠和 ABLV 的任何不良混淆,并在平衡消极态度与保护效益方面带来了任何挑战?ABLV 在澳大利亚以外的亚洲或大洋洲的其他蝙蝠种群有多远?这些问题的答案为其他地方存在狂犬病病毒提供了一个警示,在监测系统和公共卫生/兽医基础设施方面,可以更充分地认识到一种由 RABV、ABLV 及其系统群 I 病毒亲属引起的致命(尽管被忽视)疫苗可预防的疾病。
10. 蝙蝠、狂犬病病毒和血清学
所谓“无狂犬病”地区的一些流行病学家可能持有一种观点,大意是“……如果狂犬病存在的话,我会认出它……“。撇开专业意见不谈,多起流行病学事件证明并非如此。当狂犬病发生在狗等家养物种中时,人类病例通常也会随之发生,但经常被误解,尤其是在未被充分重视的易位事件之后(例如,到巴厘岛等岛屿)。如果野生动物狂犬病占主导地位,而家养物种通常没有明显的溢出感染,那么由于明显的监测偏差,这种疾病在功能上是一种消退性疾病,那么明显的“流行病学沉默”占主导地位。显然,通常的嫌疑人不会出现在鉴别诊断列表中,而这些嫌疑人被认为不是本土发生的。这种臭名昭著的先例最终在雪貂獾和 TWBLV(在一个中型岛屿上)或 ABLV(在整个大陆上)中检测到 RABV,这两者都是在公共卫生系统高于平均水平的地方。
除了单纯的临床怀疑之外,还可以通过检测抗原、扩增子或抗体进行实验室确认来对狂犬病病毒活性进行动物流行病学监测。对于后者,需要使用已知的狂犬病病毒物种之一进行中和测试(例如,FAVN、RFFIT等)或体外测试,例如ELISA(前提是有合适的分析物)。在发现手翅目动物的每个大陆上,狂犬病病例都出现在人类和家畜身上(表 1)。因此,蝙蝠是此类血清学监测研究中最常见的哺乳动物类群之一,因为与手翅目相关的大量狂犬病病毒、它们的免疫反应性以及流产性病毒感染的可能性。血清学技术很有用,因为可以安全地获得血液样本而不会对蝙蝠造成伤害。从长达数年的纵向研究来看,检测到此类抗体被解释为暴露于给定的病毒变异,并不表示存在狂犬病。由于共享的病毒表位之间存在多种具有交叉反应性的血清群,如果使用“狂犬病病毒 x”作为分析物时出现阳性蝙蝠血清,则结论不一定是存在“狂犬病病毒 x”。相反,它或相关成员负责该主机中的暴露。例如,针对 RABV 的血清会对 ABLV、DUVV 等发生交叉反应,并且不能得出 RABV 持久性是有效的结论,而不是另一个系统群成员。如果没有使用系统群 II 或 III 中的狂犬病病毒,则无法得出关于这些类群(或其他成员或等待发现的类群,因为一个受到未知限制)中成员的存在的结论。在只有一种狂犬病病毒永久存在的地理区域,监测更加简化,例如在新大陆,因为基于数十年的监测,RABV 是整个地区唯一的狂犬病病毒。然而,血清学监测在一定程度上很复杂,不仅因为需要适当的感兴趣病毒制剂,还因为可以从大多数蝙蝠类群中获得少量血清。此外,还需要阳性和阴性对照血清以及所用临界范围的标准。在全球范围内,许多研究发现阳性蝙蝠血清表明狂犬病病毒活性(补充表S5)。
这种抗体对蝙蝠狂犬病病毒的持久性可能是终生的[521]。此外,单个宿主中的抗体动力学可能因裂解病毒变体而异。例如,在尼日利亚,从为人类食用而捕获的 Eidolon helvum 果蝠中采集的样本显示出不同的中和效果,基于亚谱系:LBV-A (63%);LBV-D (49%);LBV-C (45%);和 LBV-B (24%)。这些数据表明,LBV暴露很常见。纳入各种物种分布的信息有助于考虑哪些狂犬病病毒可能在不同的类群中传播。例如,翼龙属(又名狐蝠)的广泛旧世界分布及其与 ABLV、GBLV 等的流行病学关联,为对这些和其他狂犬病病毒的血清学监测提供了可靠的理由,作为考虑在印度尼西亚、马达加斯加、马来西亚、大洋洲、东南亚等地进行流行病学反省的起点。
人类接触作为丛林肉猎杀的野生动物的比例可能更高,因为较大的翼龙在某些文化中受到重视,而且它们的神秘特性被某些文化所重视,例如在澳大利亚的部分地区。甚至较小的物种也可能被纳入部落节日,例如在非洲部分地区。用于潜在疾病生态学考虑的其他广泛分类群可能仅包括那些分布广泛且丰富(即非濒危、非致命血清学采样方法最不受关注的状态)的物种代表 Eidolon、Epomophorus、Eptesicus、Miniopterus、Myotis、Nycteris、Rousettus 等。
当缺乏对家畜或野生动物(如蝙蝠)进行狂犬病病毒检测的常规监测活动时,风险建模是另一种方法,对于那些认为自己“无狂犬病”但缺乏基于实验室的验证标准的地方来说,可以考虑优先考虑。由于血清学检测提供的病毒活性证据比受影响人群中抗原或扩增子的检测高出几个数量级(即,每个样本数量<1/100–1/1000 个阳性),样本量至少为 ~30 个个体(基于全球发现的血清阳性率),通过对高丰度物种的纵向研究积累的此类数据为长期生态监测提供了合理的考虑。
11. 对狂犬病当前和未来的担忧
除南极洲外,狂犬病病毒广泛分布于所有大陆(但鉴于最近毛皮海豹爆发了大规模疫情,即使是该大陆也面临着多种海洋哺乳动物潜在感染的风险)。这些不同的 RNA 病毒都会引起急性进行性脑炎,通常通过受感染的狂犬病哺乳动物的叮咬传播(尽管鸟类也易感)。尽管历史上被认可并作为“同一健康”优先事项进行了相关范例,但这种独特的人畜共患病继续给农业、公共卫生和保护生物学带来巨大负担。此外,当“病因”和“疾病”之间的历史差异没有得到重视时,就会出现病毒学混淆。可以理解的是,如果调查人员将专注于局部控制、区域预防或选择性消除的流行病学术语误认为是真正的“根除”,学术界就会感到震惊。同样,如果某些政治单位认为自己“没有疾病”,但一些客观实验室监测的国际标准没有得到满足,特别是面对可能的野生动物宿主的神秘存在,也可能会出现挑战。自 19 世纪以来对相关病理生物学有基本的了解,自 Negri/Williams 以来存在改进的灵敏和特异性诊断测试,以及从 1920 年代至今越来越安全和有效的生物制剂的可用性,这些病原体和这种疾病仍然被忽视,即使有 20 世纪工具的谨慎影响,这似乎相当奇怪。这不仅仅是对中低收入国家内部情况的关注,对美国近期监测数据的简要反思说明了这种担忧。
在控制和消除犬狂犬病之前,大多数人类病例是由患有狂犬病的狗引起的。如今,美国的人类病例很少见,通常与蝙蝠RABV感染有关,或由于境外暴露的输入病例。2019年和2020年均未报告人类狂犬病病例,而2021年则有5例死亡。此后,2022年和2023年均未报告人类狂犬病死亡病例。然而,到 2024 年,报告了 5 例病例:东北狭长地带地区疑似人类狂犬病死亡狂犬病病毒一名加州教师在教室里被蝙蝠咬伤;明尼苏达州一名老年妇女暴露于蝙蝠;肯塔基州一名据信在加勒比地区暴露的人;以及一名于 12 月死亡的人,他是密歇根州一名移植受者的器官捐献者,随后于 2025 年去世。这样一个名副其实的流行病学脚注揭示了一个高度发达、无狗狂犬病的国家罕见事件的生物学,以及何时、何地、如何或为何出现狂犬病病毒感染的不可预测性。
12. 结论
我们已经证明,人畜共患病狂犬病,如一些系统群 I 狂犬病病毒所说明的那样,适合控制、预防或选择性消除(即某些病毒物种的少数变体),但这种疾病不会被根除。相反,对于由多种相关病原体引起的同一疾病,我们非常谨慎,我们强烈建议所有兽医专业人员: 区分病因和疾病;接受 PrEP;向任何有接触风险的家畜接种经批准的疫苗;了解狂犬病病毒带来的公共卫生问题(以及该疾病无法治愈);识别疑似患者的临床症状;约束、镇静、安乐死并收集适当的脑组织进行测试;遵守生物安全实践;使用个人防护装备;报告阳性病例结果以进行必要的通知;向任何可能接触的客户提供咨询,说明需要及时就医,包括考虑 PEP;欣赏一些动物水库目前还不为人知;认为如果没有相当客观的诊断性内省,地方实际上可能不会“没有”“自然病”;并鼓励国家当局将病原体发现和病毒表征作为现代实验室监测的常规部分。
此外,从兽医病毒学的基本角度来看,并非一切都能解决。到 2025 年,哺乳动物宿主的广度没有已知的限制,根据其现存的近亲,对 RABV 最近的病毒学“成功”没有深入的基本理解,没有泛瑞莎病毒生物制剂,没有获得许可的抗病毒药物,没有即将出现的通用疗法,也没有学术共识是否会很快出现实质性宿主转移或何时会欣赏“新”狂犬病病毒。在如此明显的限制下,相比之下,目前消除犬类狂犬病的全球路线图和接近“ZBT”大幅改进的时间表似乎几乎很简单,这构成了真正的跨学科挑战。
Hits: 933
- 狂犬病(2025年6月更新)
- 狂犬病 你应该知道的
- 预防和治疗狂犬病的狂犬病疫苗
- 狂犬病可能成为历史
- 看看每种疫苗:狂犬病疫苗
- 阿拉巴马州妇女在从车上卸下杂货时被狂犬病狐狸咬伤:“公共卫生威胁”
- 应对狂犬病
- 全球疫苗免疫联盟将促进全球疫苗免疫联盟、世卫组织和阿联酋50多个国家获得拯救生命的人类狂犬病疫苗
- 美国CDC发布了将狗带入美国的新规定,旨在防止狂犬病
- 全国狂犬病监测方案
- 2024年东帝汶首例狂犬病确诊病例
- WHO实况报道 狂犬病
- 狂犬病
- 狂犬病疫苗接种:给医疗保健提供者的信息
- 特定感染的流行病学和控制:狂犬病
- 狂犬病疫苗接种的注意事项或禁忌症
- ACIP 使用减少(4剂)疫苗计划进行暴露后预防以预防人类狂犬病
- 实施良好的狂犬病暴露后预防方案
- ACIP 使用修改后的暴露前预防疫苗接种计划预防人类狂犬病,美国,2022年
- 西班牙犬类狂犬病免疫保护的影响因素
- 减少犬的进口等待期-对狂犬病恐惧科学合理的变化
- 老年人对狂犬病暴露后预防无应答:挑战和方法
- 狂犬病疫苗的效力、免疫原性和有效性:体外和体内方法
- 狂犬病人免疫球蛋白 (HRIG)
- “震惊”的妇女,超过6,000美元的救命狂犬病治疗账单
- 负鼠感染狂犬病毒对这种病毒在城市环境中的传播敲响了警钟
- 索马里蜜獾感染未报告的首例疑似人狂犬病病例研究
- 两名狂犬病研究人员解释如何保护自己免受这种疾病的侵害
- 临床医生常见的狂犬病问题
- 狂犬病疫苗
- 世界卫生组织关于狂犬病的常见问题
- 狂犬病实况报道
- 狂犬病
- 狂犬病是一种古老的、不可预测的、可能致命的疾病——两位狂犬病研究人员解释了如何保护自己
- 狂犬病疫苗短缺:给卫生专业人员的建议
- 7例被狂犬咬伤者接种狂犬病疫苗的效果观察
- 全球狂犬病风险:狂犬病疫苗短缺期间的实用建议
- 巴西一个土著村庄被蝙蝠掠夺的高风险:警告狂犬病在马卡利人中重新出现
- 评估在澳大利亚蝙蝠狂犬病病毒和狂犬病暴露后预防中血清学的益处
- 蝙蝠处理者的狂犬病暴露前预防
- 基于狂犬病中和抗体水平的狂犬病加强剂时机指南
- 施用狂犬病疫苗和免疫球蛋白
- 为狂犬病暴露后预防开发的单克隆抗体鸡尾酒疗法
- 狂犬病预防处置常见问题解答(成大生物)
- 世界卫生组织狂犬病专家磋商会 第三版报告
- 世卫组织对狂犬病的看法
- 新型狂犬病疫苗
- 狂犬病病毒可以抗强大的天然免疫
- 狂犬病的被动免疫
- 亚洲狂犬病预防
- 秘鲁高风险人群的大规模狂犬病暴露前预防
- 人用狂犬病疫苗
- 野生动物狂犬病疫苗
- 狂犬病和狂犬病疫苗的历史
- 狂犬病的生命周期、传播和发病机制
- 狂犬病暴露后管理指南(英国 2023年1月)
- 2022年印度马哈拉施特拉邦一名幼儿出现临床狂犬病症状后存活
- 第27章:狂犬病
- 狂犬病:问题和答案
- 《狂犬病暴露预防处置工作规范(2023年版)》解读问答
- 狂犬病暴露预防处置工作规范(2023年版)
- 狂犬病
- 狂犬病相关问题解答
- 疾病控制中心加强了对进口狗的狂犬病管理,原因如下
- 关于狂犬病
- 一名加州艺术教师在教室遭遇蝙蝠后死于狂犬病
- WHO 实况报道 动物咬伤 2024年1月
- 疫苗接种和免疫 狂犬病
- 世卫组织推荐皮内途径用于狂犬病暴露后预防
- 狂犬病疫苗:世卫组织立场文件–2018年4月
- WHO实况报道 狂犬病 2024年6月
- 狂犬病-结束忽视, 实现可持续发展目标 被忽视的热带病路线图 2021-2030
- 动物狂犬病
- 宠物主人对疫苗的犹豫越来越多——一位公共卫生专家解释了为什么这很重要
- 兽用狂犬病疫苗
- 何时寻求狂犬病医疗护理
- 猫
- 关于获得人类狂犬病疫苗的社会正义观点
- 评估新的狂犬病暴露后预防(PEP)方案或疫苗
- 卫生和福利部-狂犬病
- 狂犬病暴露后预防用单克隆抗体开发的最新进展:临床开发管线现状综述
- 世卫组织对狂犬病免疫接种的立场 – 2018 年最新情况
- 恐水症:1874 年的狂犬病和改变一切的救命疫苗
- 新的人类狂犬病疫苗正在研发中
- 为什么消除狂犬病?
- 为什么现在要根除狂犬病?
- 狂犬病-NFID
- 为狗接种疫苗 – 它们不会因接种疫苗而患上自闭症
- 宠物主人版 狗的狂犬病
- 猫的狂犬病
- 狂犬病-国际脑炎协会
- 器官移植后如何感染狂犬病?
- 器官移植患者感染狂犬病后死亡
- 人类狂犬病疾病负担:Rabipur®疫苗的潜在预防作用
- 印度东北部农村的狂犬病:强调同一健康方法紧迫性的病例报告
- 狂犬病毒糖蛋白的深度突变扫描确定了突变限制和抗体逃逸突变
- 人类狂犬病:一种治疗方法
- 通过受感染狗唾液污染的隐形眼镜传播狂犬病病毒(病例报告)
- 越南暴露前预防计划中基于灭活 vero 细胞的狂犬病疫苗 (Rabivax-S) 的安全性和免疫原性
- 狂犬病生物制剂在 III 类动物暴露儿科患者中的安全性和临床疗效的前瞻性研究
- 重复狂犬病疫苗接种揭示人类中和抗体的分子趋同
- 狂犬病病毒的感染和预防
- 关于 Zagreb 或 Essen 方案后加强疫苗接种的免疫原性和安全性的 3 期临床试验
- 通过 RABV-G 结构域筛选和异源初免-加强免疫优化狂犬病 mRNA 疫苗功效
- 狂犬病实验室诊断的最新情况
- 三色蝠相关狂犬病毒引起的一例人狂犬病的报道
- 一种核苷修饰的狂犬病mRNA疫苗在小鼠和非人灵长类动物中诱导出持久而全面的免疫反应
- 狂犬病的识别和治疗:病例报告和讨论
- 人类狂犬病的诊断困难:病例报告和文献回顾
- 狂犬病感染的边缘系统症状
- 狂犬病脑炎的生存率
- 人类狂犬病:2016年更新
- 突尼斯一名狂犬病幸存儿童:病例报告
- 肯尼亚蝙蝠携带的Duvenhage病毒导致人类狂犬病死亡:昏迷诱导、氯胺酮和抗病毒药物治疗无效
- 病例报告:狂犬病幸存者长期随访中的神经功能恢复与血清学反应
- 蝙蝠和食肉动物中狂犬病的比较病理学及其对人类溢出影响
- 病例报告:狂犬病生存者:印度病例系列
- 评估人类抗狂犬病疫苗的反应:一种快速、高通量适应性、基于假病毒的中和试验,作为快速荧光病灶抑制试验 (RFFIT) 的替代方案
- 病例报告:狂犬病幸存者长期随访中的神经功能恢复与血清学反应
- 巴西巴伊亚州狂犬病爆发的流行病学与分子分析
- 俄罗斯下诺夫哥罗德地区动物狂犬病(2012-2024年):描述性与预测性流行病学分析
- 巴西东南部一只黑鼠耳蝠(黑鼠耳蝠属物种Myotis nigricans)感染狂犬病病毒、科勒巴尔通体菌以及婴儿利什曼原虫的混合感染情况
- 2013-2023 年悉尼大都会居民狂犬病暴露后治疗:回顾性病例系列分析
- 用肌肉靶向脂质纳米颗粒配制的狂犬病病毒 G 蛋白 mRNA 在小鼠体内的免疫原性
- 关于狂犬病的 One Health Lens:马拉维利隆圭农村狂犬病病毒的人类-蝙蝠相互作用和基因组见解
- 阿根廷低地貘 (Tapirus terrestris) 麻痹狂犬病的首次报告
- 由表达狂犬病糖蛋白的非复制型病毒载体痘苗病毒天坛株改良株(MVA)和腺病毒5型(Ad5)所诱导产生的对狂犬病的保护作用
- 印度野生猫科动物中狂犬病毒的免疫病理与分子筛查:揭示从野外到城市的溢出效应
- 韩国貉 (Nyctereutes procynoide koreensis) 接种了含有 ERAGS-GFP 毒株的新型狂犬病诱饵疫苗的保护性抗体反应
- 宠物护理 疫苗
- 为您的宠物接种疫苗
- 狂犬病感染:如何保护您的孩子
- 新型狂犬病暴露后预防单克隆抗体鸡尾酒疗法的安全性和耐受性
- “愤怒的狂犬病”人类图画案例
- 蝙蝠咬伤
- 研究蝙蝠病毒如何帮助预防人畜共患疾病
- 狂犬病:症状繁多、宿主广泛的恶魔病毒——罗德尼·罗德解读
- 狂犬病的同一健康:不仅适用于动物
- 狂犬病和蛔虫:浣熊相关传染病
- 丽莎病毒 很罕见,但致命。如果蝙蝠咬你怎么办?
- 海地在四个省开展新的疫苗接种运动,加强防治犬类狂犬病
- 家养及其他动物狂犬病病毒感染的挑战:病毒学层面的困惑、惊愕、懊恼与好奇交织
- 双重战斗:狂犬病和蛇咬伤在东南亚的毒害
- 狂犬病在城市无人管理的猫群中爆发 — 马里兰州,2024 年 8 月
- 尼泊尔正力争在2030年前消灭狂犬病,但领导人表示,现在是加大疫苗投资的时候了。
- 现代狂犬病疫苗的诞生历程
- 一种基于mRNA的新型狂犬病病毒多表位疫苗:通过反向疫苗学和免疫信息学进行计算机设计
- 快速荧光灶抑制试验(RFFIT)的验证与评价 —— 作为狂犬病疫苗相对效价间接检测方法的中和抗体定量检测:NIH 试验的替代方案
- 暴露后狂犬病预防失败案例研究
- 2021年明尼苏达州一起经确认的人畜共患病暴露后,疑似因宿主介导导致暴露后预防失败的致命人类狂犬病感染病例
- 给野生动物投喂疫苗:口服狂犬病疫苗的故事
- 降低肯尼亚西部臭名昭著的狗市狂犬病风险
- 印度的狂犬病
- 海地的狂犬病
- 2024年10月至2025年2月,美国发生通过实体器官移植传播狂犬病的病例,该器官移植供体未确诊患有狂犬病
- 两例死亡与狂犬病感染的肾移植有关
- 明尼苏达州一家奶牛场的牛群中爆发狂犬病聚集性疫情——2024年
- 摩洛哥的狂犬病
- 2024年肯塔基州和俄亥俄州输入性人类狂犬病病例
- 2024年明尼苏达州和加利福尼亚州人类狂犬病死亡人数
- 反疫苗人士对宠物狂犬病 RNA 疫苗的虚假宣传
- 新型口服狂犬疫苗能否帮助我们最终终结这种疾病?
- NACI 2026 年 1 月 30 日发布的声明摘要:狂犬疫苗用于暴露前预防 (PrEP) 的最新指南
- 狂犬疫苗的给药途径:数据比较
- 猫会得狂犬病吗?
- 2022-2024年中欧狂犬病疫情再度出现
- 研讨会报告:改进狂犬病血清学方法和程序的标准化
莫博士请问一下我们显示的评论,但是你没回复的是不是就是指我们肯定没有问题?,,祝您休假开心万分,感谢孟博士,
还有事我不知道
孟博士,请教您我加强针今年四月份打的,但是总是买蔬果什么的总是担心狗舔舐过,这个是不是对我来说肯定没问题呢,谢谢孟博士,祝您休假开心,万分感谢!
孟博士好,狂犬病毒会复制吗?沾有狂犬病毒的衣物,或是物品,在没有清洗消毒和阳光暴晒的情况下,碰到了其他的物品,其他的物品或是皮肤上会沾上狂犬病毒吗?新鲜伤口接触这些被流浪动物唾液污染的物品衣物,会感染狂犬病毒吗
孟教授,狗一个月没死,请问狗还有传播狂犬病的风险吗?
你好,孟博士,我是华,前面加强的时间不记得了,之后我记得是23年3月加强两针,23年11月加强两针,24年7月加强过两针,
我今年25年7月8号暴露了,暴露延迟10(7.18)又去加强了两针,我打完第一加强针三天过10个小时后抽血送去了中抗广州实验室测抗体,结果是33.83。我就想问有没有可能我打加强针前体内抗体低于0.5。因为我看过您的博文介绍,打完加强针三天抗体变化不大,可我是打完第一针加强针过三天10个小时才测的,问题如下:
1.会不会多这个10小时把我抗体从低于0.5突然暴涨到30多?
2.还是说我7.18号前实际抗体浓度还是高于0.5的?
因为网上有关这方面的说法到处都有,没有人能准确给我解释,所以我很纠结。
辛苦孟老师有时间回复一下。
孟博士您好,1.去年7.3号打完暴露前预防,去年8.23号测试抗体4.88,去年12.11号测试3.68,到今天抗体还是高于0.5抗体如果这个时候发生暴露需要加强吗
2.这种抗体变化正常么?
3.您觉得按照这种抗体变化情况三针狂犬疫苗保护两年应该没问题吧?
大于0.5可以不加强,但如果是高危人群,建议低于1就加强
孟博士您好,阅读您之前的博文后感触很深,目前感觉已经脱恐。我试图总结了一些通用的观点,希望能帮助到更多的人,不知道我下面的四条总结的是否正确,想找您确认下,恳请孟博士批评指正:
1. 虽然过去可能发生“狂犬疫苗接种过量、接种不规范、有时暴露后未去接种、服药、生活不规律”等各种复杂情况,但是只要过去接种过狂犬疫苗,目前未发病(狂犬病)且目前测得狂犬病毒中和抗体>0.5IU/ML,就可以证明本次测抗体之前所有发生及可能发生的暴露所带来的狂犬病风险都不存在,不需要担心感染狂犬病。(注:如果测抗体时精神状态正常,则极大概率没有发病;如发病了,很快就会有狂犬病症状并且极大概率短时间内死亡。)
2. 虽然过去可能发生“狂犬疫苗接种过量、接种不规范、有时暴露后未去接种、服药、生活不规律”等各种复杂情况,但是只要过去接种过狂犬疫苗,就可以使用下述测抗体的脱恐策略(即每半年测一次狂犬病毒中和抗体,如果大于0.5IU/ML,不用做任何处理;如果小于0.5IU/ML,可以考虑加强一针。逐步降低检测频次,直到脱恐。注:如果期间发生明确暴露,请及时就医)
3. 对于恐狂症患者,在发生明确暴露的情况下,及时就医;在没有发生明确暴露的情况下,减少臆想,多锻炼,增强自己的判断能力,让自己相信自己的眼睛、记忆。
4. 对于恐狂症患者,要增加对极小概率事件的接受能力,极小概率事件在实际生活中本身就没有关注的必要,过于纠结极小概率事件往往也得不到答案。
毛线,总结这么牛
感谢孟博士的鼓励和肯定!希望能帮到更多的人!
孟博士,去年7.3号打完暴露前预防,去年8.23号测试抗体4.88,去年12.11号测试3.68,到今天抗体还是高于0.5抗体如果这个时候发生暴露需要加强吗?还有就是这种抗体变化正常么?三针效果感觉能保护两年啊
1.没有暴露不加强
2.确信大于0.5,也不用加强
3,有些厂家疫苗可以的
谢谢
孟老师,请问您除了器官移植以外与病人接触会感染吗?只是正常接触,没有一起吃饭之类的,打了加强针是不是就没事了呢?没得到您的回复我实在是害怕,因为我亲戚最近发烧而且他被狗咬过还没打疫苗,还有十多天到一年,万分感谢孟老师您的回复,因为我一得到您的回复马上就不害怕了,因为您和严老师是这方面专家,,感谢孟老师您,
其它未报到
孟博士,您好!想请教一个问题,平时在公共食堂或者路边摊吃饭,因为经常有流浪狗出现,会存在有人故意在饭菜中下狂犬病毒吗,如果吃了这样的饭菜有感染风险吗,感谢孟博士的解答,祝孟博士身体健康,事事顺利。
故意???
孟博士,你好!是不是认为没有这种传染方式的报道啊
孟博士你好 我是那个测出抗体33.83的人 今天距离上次抽血第10天了 感觉今天喝水喉咙已经开始痉挛了 我也没有出现什么发热前驱期 不知道为什么会这样
毛线;这么高的抗体,还担心啥呢
孟博士您好,我去年7.3号打完了三针狂犬疫苗,然后到12月份还有3点多抗体,这个月7号我不小心把自己腿部挠破我去厕所冲洗后出来,在我家阳台感觉皮肤破损处附近被水溅到了一下,我家阳台露天有玻璃但是玻璃附近是防虫网,我就担心会不会是蝙蝠口水,这种情况需要担心吗?今天突然腿一直用力抬起来一会后发酸把我吓得还用打加强针么?
你已经开始担心了,我从未听说这中传播方式
那还需要打加强针吗?
孟博士您好,以前问问题您回复我,瞬间我就不恐了,不知道怎么感谢您,就博士书房看到文章会默默的点赞!
今天胳膊划了2道红印,刚才我去一个养狗的农村亲戚家里,没等下车就已经把狗关起来了,我在他家待了会,没看到狗,但是看到隔壁的猫了,亲戚就拿石头给它打跑了,但是临走时候我发现了胳膊2个红印,咋整的也想不起来了,我在4.28和6.23打了2针生物制剂,请问我去年10月24打的加强第二针,还有2个多月就一年了,如果是猫狗划的我安全吗?
公众号不是回复了吗